

Биоэлектрические потенциалы
Биоэлектрические потенциалы (биотоки) — электрические явления, наблюдаемые в живых клетках в покое и при физиологической деятельности. Возникновение в живых клетках электрических потенциалов и обусловленных ими биотоков связано с физико-химическими свойствами клеточных мембран и компонентов цитоплазмы (аминокислот, белков, ионов). Между наружной поверхностью клеточной мембраны и внутренним содержимым клетки существует всегда разность потенциалов, которая создается в силу различной концентрации ионов К+, Na+, Cl- внутри и вне клетки и различной проницаемости для них клеточной мембраны. Эта разность потенциалов называется «током покоя», или мембранным потенциалом, и составляет в среднем 60—90 мВ. При возбуждении живой клетки происходят изменения исходного мембранного потенциала за счет изменения проницаемости мембраны и перемещения ионов. В клетках возбудимых тканей (мышечной, нервной) эти процессы могут происходить в очень короткие интервалы времени (миллисекунды) и называются «током действия». Величина его может достигать 120 мВ. Для отведения биотоков от отдельных клеток организма попользуют специальные стеклянные и металлические микроэлектроды, имеющие неизолированные кончики (1 — 2 мк). Регистрация биоэлектрических потенциалов осуществляется с помощью катодных осциллографов и различных чернильнопишущих устройств с использованием усилителей биопотенциалов. См. также Электроэнцефалография, Электрокардиография, Электромиография,Электроретинография.
Биоэлектрические явления (биоэлектрические потенциалы, биотоки) — электрические процессы, характерные для живых тканей. Биоэлектрические явления открыты Гальвани (A. L. Galvani) и Маттеуччи (С. Matteucci). Первые гипотезы о природе биоэлектрических явлений выдвинуты Дюбуа-Реймоном (Е. Du Bois-Reymond) и Германом (L. Hermann). В. Ю. Чаговец разработал теорию биоэлектрических явлений на основе современной физической химии электролитов. Подробное изучение биоэлектрических явлений стало возможным лишь с созданием соответствующей измерительной аппаратуры (катодные и шлейфные осциллографы в сочетании с высокочувствительными электронными усилителями). В настоящее время разработаны методы, позволяющие вводить электроды в глубь отдельных клеток живой ткани и регистрировать их электрическую активность. Такие исследования показывают, что между наружной и внутренней поверхностями клеточной мембраны существует постоянная разность потенциалов, причем наружная поверхность по отношению к внутренней имеет положительный заряд. Величина такой «трансмембранной» разности потенциалов составляет несколько десятков милливольт; она обозначается как «мембранный потенциал» или «потенциал покоя». Изменения потенциала покоя при различных функциональных состояниях клетки имеют названия: «потенциалы действия», «синаптические потенциалы», «генераторные потенциалы», «секреторные потенциалы» и т. д. Основой постоянной электрической поляризации поверхностной мембраны клетки является неравномерное распределение неорганических ионов (в первую очередь калия, натрия и хлора) между протоплазмой клетки и ее средой. В протоплазме клетки имеется значительный избыток ионов калия при сравнительно небольшом количестве ионов натрия и хлора (так называемая ионная асимметрия). В покоящемся состоянии клетки распределение ионов стационарно и поддерживается при неодинаковой проницаемости клеточной мембраны к различным ионам деятельностью активных протоплазматических механизмов, откачивающих определенные ионы из клетки или, наоборот, втягивающих их внутрь нее,— так называемый калиево-натриевый «насос», или «помпа». Подробно механизм деятельности этого «насоса» пока не выяснен, однако установлено, что он связан с источниками метаболической энергии клетки, особенно с системой расщепления макроэргич. фосфорных соединений. Ионы натрия и хлора оказывают влияние на потенциал покоя лишь при низких концентрациях ионов калия вне клетки. Наиболее точное определение величины разности потенциалов, существующей на клеточной поверхности, может быть проведено микроэлектродным методом, когда один из отводящих электродов (стеклянная микропипетка с диаметром кончика менее 0,5 мк) введен внутрь клетки, а вторым электродом служит окружающая клетку ткань. У млекопитающих, например, потенциал покоя нервной клетки составляет 60—80 мВ, мышечного поперечнополосатого волокна — 80—90 мВ, сердечного — 90—95 мВ. При возбуждении клетки изменяется ионная проницаемость ее поверхностной мембраны, вследствие чего появляются кратковременные перемещения ионов через мембрану. Эти ионные токи являются причиной изменений электрической поляризации мембраны клетки. Механизм возникновения наиболее распространенной формы активной электрической реакции — потенциала действия, связанного с распространяющейся волной возбуждения,— можно представить следующим образом. Необходимым условием возникновения распространяющегося возбуждения является снижение величины потенциала покоя (деполяризация) до определенной величины (истинный порог возбуждения клетки). Механизм возбуждения деполяризацией мембраны универсален; такое возбуждение возникает не только при электрическом, но и при любых других видах раздражения, в том числе при адекватных раздражениях рецепторных окончаний. Когда деполяризация достигает критического уровня (различного для разных типов клеток), стремительно развивается кратковременное повышение ионной проницаемости клеточной мембраны для таких ионов, которые в покоящемся состоянии с трудом проходят через мембрану. Природа этих изменений неизвестна; установлено, что в большинстве случаев ионом, который начинает особенно легко проходить через клеточную поверхность, является натрий. Положительные заряды ионов, движущихся внутрь клетки, не только полностью устраняют потенциал покоя, но даже на короткий момент так извращают трансмембранную разность потенциалов, что наружная поверхность мембраны становится отрицательной по отношению к внутренней ее стороне. В итоге этих процессов на поверхности клетки создается продольная разность потенциалов — возбужденный ее участок оказывается отрицательно заряженным по отношению к невозбужденному. Возникающие в связи с этим кольцевые электрические (ионные) токи между участком возбуждения и соседними невозбужденными участками («токи действия») являются причиной деполяризации невозбужденных участков до порогового уровня, что и обеспечивает распространение волны возбуждения по клетке. Амплитуда потенциала действия и его длительность у нервной клетки млекопитающих соответственно составляют 100—110 мВ и 1—2 мсек, у поперечнополосатого мышечного волокна — 110—120 мВ и 3—5 мсек. Однако потенциал действия у сердечного мышечного волокна оказывается чрезвычайно длительным. У каждой клетки амплитуда потенциала действия в нормальных условиях постоянна независимо от условий раздражения (правило «все или ничего»); однако если потенциал покоя по каким-либо причинам чрезвычайно снижен, то и амплитуда потенциала действия начинает уменьшаться либо генерация последнего оказывается совершенно невозможной («катодическая депрессия» или «инактивация»). После окончания основной «высоковольтной» части потенциала действия (пик) происходят дополнительные незначительные по амплитуде колебания электрической поляризации клеточной поверхности (так называемые следовые потенциалы). Выраженность их весьма варьирует у различных типов клеток. В большинстве случаев сразу после пика развивается следовая деполяризация, сменяющаяся следовой гиперполяризацией, которая, например, в соме нервной клетки достигает 100 мсек. Развитию процесса возбуждения соответствует и определенная теплопродукция, которая точно изучена на изолированном нерве и составляет в I стадии (начальное теплообразование) 7·10-8 кал на 1 г нерва на импульс, а во II стадии (отсроченное теплообразование) значительно превышает эту величину. В большинстве случаев развитию потенциала действия предшествуют промежуточные формы электрической активности. Одной из таких форм является локальный (местный) потенциал, возникающий при околопороговых раздражениях и отличающийся от потенциала действия градуальной зависимостью от силы раздражения (т. е. от величины вызывающей его деполяризации клеточной мембраны). В случае синаптической передачи возбуждения возникает особая форма локального потенциала — постсинаптический потенциал (ПСП), который также характеризуется градуальностью и способностью суммироваться с другими аналогичными потенциалами. ПСП является результатом специфической реакции постсинаптической мембраны клетки (т. е. той части ее мембраны, к которой прилегают синаптические окончания аксонов других клеток) на действие выделяемого синаптическими окончаниями медиатора. В зависимости от характера этого медиатора, а также, по-видимому, и от особенностей тех рецепторных группировок постсинаптической мембраны, которые с ним реагируют, ПСП могут выражаться изменениями электрической поляризации мембраны в различных направлениях. В одних случаях мембрана деполяризуется; если деполяризация достигает порогового значения, то генерируется обычный потенциал действия. Такие ПСП являются возбуждающими (ВПСП) и связаны с деятельностью особых возбуждающих синаптических окончаний. В других случаях электрическая поляризация мембраны увеличивается, а возникновение потенциала действия соответственно затрудняется; такие ПСП оказываются тормозящими (ТПСП) и лежат в основе синаптического торможения. При адекватном возбуждении нервных окончаний в рецепторах внешняя энергия трансформируется первоначально в градуальную деполяризацию их мембраны (генераторный потенциал), которая уже непосредственно приводит к развитию распространяющихся нервных импульсов, если раздражение достигает пороговой величины. Особенности генераторных потенциалов хорошо изучены на механорецепторах, простейших фоторецепторах и др. Секреторные процессы также связаны с возникновением на клеточной мембране железистой клетки градуальных секреторных потенциалов, которые могут иметь различные направление и длительность в зависимости от характера секреторной деятельности.
Биоэлектрические потенциалы отдельных клеток могут суммироваться в электрические реакции целой ткани или органа. Так как ткань является проводником электричества (второго рода), то эти реакции могут быть зарегистрированы даже при расположении отводящих электродов на некотором расстоянии от органа (например, на коже). В нормальных условиях потенциал покоя клеток не обнаруживается в неповрежденной ткани, так как наружная поверхность каждой из них является изопотенциальной и имеет разность потенциалов только по отношению к внутреннему содержимому клетки. Однако если участок ткани повредить, разрушив тем или иным образом оболочки части клеток, то этот «альтерированный» участок во всех случаях окажется отрицательно заряженным по отношению к неповрежденным участкам той же ткани. Если достаточно большое количество клеток исследуемой ткани возбуждается одновременно и генерирует потенциалы действия или другие электрические реакции, то соответствующая электрическая реакция может быть зарегистрирована от целой ткани (нерва, мышцы, железы, участка мозга и т. д.). Как при повреждении, так и при возбуждении амплитуда колебаний будет значительно меньше, чем на мембране отдельной клетки, в связи с шунтированием внеклеточных токов межклеточной жидкостью. Во всех случаях электрод, соприкасающийся с возбужденным участком ткани, будет отрицательным по отношению к электроду, расположенному на невозбужденной части. Если импульс возбуждения поочередно проходит под одним и другим отводящими электродами, то будет зарегистрирован двухфазный ток действия; если же второй электрод расположить так, чтобы волна возбуждения не могла его достичь, то зарегистрированное колебание окажется однофазным. В неоднородной по клеточному составу ткани возникают более сложные типы электрических реакций, которые могут состоять из большого количества колебаний. Таким является, например, потенциал действия нервного ствола, содержащего волокна с различной скоростью проведения, если его отвести на некотором удалении от места раздражения, когда нервные импульсы в различных типах волокон уже успевают значительно разойтись во времени. От очень сложных по структуре образований, например от ЦНС, электрические колебания отводятся все время, даже при отсутствии специальных раздражений. Это связано с непрерывным поступлением импульсации от большого количества неконтролируемых источников, возможностью длительной циркуляции нервных импульсов по сложным цепям нейронов, а также генерацией ритмических импульсов нервными клетками под влиянием химических и других факторов. Электрическая активность соответствующих участков мозга проявляется ритмическими колебаниями очень небольшой амплитуды (несколько десятков мкв), отражающими общее функциональное состояние нейронов в соответствующей области мозга (хотя механизмы появления ритмических колебаний потенциала и изменения ритма при различных функциональных состояниях мозга пока не ясны). Биотоки, отведенные от поверхности коры больших полушарий, получили название электроэнцефалограммы; их регистрируют и изучают для физиологических и клинических исследований деятельности головного мозга (см. Электроэнцефалография). При раздражении различных сенсорных систем (рецепторы, афферентные нервы) в определенных областях мозга регистрируются характерные биоэлектрические ответы (так называемые вызванные потенциалы). Они имеют четкую локализацию на различных уровнях ЦНС и отражают некоторые характерные параметры возбудимых систем (латентный период). Путем регистраций вызванных потенциалов изучают пути передачи сенсорных импульсаций и их кортикальные проекции (локализацию). Широкое применение нашла регистрация суммарных электрических реакций сердечной мышцы (электрокардиография), скелетной мускулатуры (электромиография), сетчатой оболочки (электроретинография).
Биоэлектрические явления
Биоэлектрические явления подробно рассматриваются в курсе электрофизиологии, однако следует отметить, что изучение сущности этих явлений еще далеко не закончено. [c.53] В пособии (первое издание в 1969 г.) излагаются основные современныезнания о возрастных особенностях строения и функциях организма детей дошкольного, младшего и среднего школьных возрастов, излагаются новейшие материалы об индивидуальном развитии человека и о современной акцелерации развития. Строение клеток дается на основе электронномикроскопических исследований. Биоэлектрические явления рассматриваются в свете мембранной теории. Представлены современные сведения о передаче нервного импульса,природе торможения, развитии головного мозга у детей, особенностях высшейнервной деятельности в связи с формированием их психики и задачами педагогики — обучения и воспитания. Приводятся основные сведения, необходимые дляорганизации труда детей и их физического воспитания.. [c.143] Бурлакова Е. В., Вепринцев Б. H., Колье О. Р., Кригер Ю. А., Исследования биоэлектрических явлений в тканях и клетках. Изд. Высшая школа , 1961. [c.172] Современный этап развития биофизики начался, по существу, с выдающихся открытий Л. Полингом пространственной структуры белка и Д. Уотсоном и Ф. Криком знаменитой спирали жизни - двойной спирали ДНК. Последовательноеприменение физических методов и представлений при изучении надмолекулярных мембранных структур привело к открытию ионной природы биоэлектрических явлений (А. Ходжкин, А. Хаксли, [c.5] БИОЭЛЕКТРИЧЕСКИЕ ЯВЛЕНИЯ В НЕРВНЫХ КЛЕТКАХ [c.246] Кальций выполняет многообразные функции в обмене веществ клеток и организма в целом. Они связаны с его влиянием на структуру мембран, ионные потоки через них и биоэлектрические явления, на перестройки цитоскелета,процессы поляризации клеток и тканей и др. [c.248] Богуславский Л. И. Биоэлектрические явления и граница раздела фаз.—М. Наука, 1978.— 360 с. [c.299] Замечательным результатом такого подхода являются ионоселективные электроды. Их развитие может быть прослежено от исследования природныхбиоэлектрических явлений до создания искусственных мембранных систем и — вобратном направлении — от создания мембранных систем к попыткам объяснения биологических процессов на клеточном уровне. [c.11] В 1848 г. Дюбуа-Реймон [21] высказал мысль, что поверхности биологических структур имеют общее свойство с электродом в гальваническом элементе, и именно это свойство является причиной биоэлектрических явлений, наблюдаемых в поврежденных тканях. Свойства биологических мембран не могли быть, однако, объяснены до тех пор, пока не были сформулированы покрайней мере основные положения электрохимии простых мембранных моделей. Термодинамические зависимости для мембранных равновесий былиполучены Гиббсом в 1875 г. [29]. Однако, поскольку теория растворовэлектролитов впервые была [c.11] К числу наиболее удивительных свойств живых систем несомненно следует отнести их способность генерировать биоэлектрические потенциалы. Обнаруженная в середине прошлого столетия, она впоследствии привлекла к себе внимание исследователей самых разных направлений биологов, физиков, физико-химиков и др. В результате изучение биоэлектрических явлений приобрело многоплановый характер. Одним из существенных направлений, получивших развитие, явилось изучение биоэлектрогенеза. т.е. непосредственных причин, лежащих в основе генерации биоэлектрических потенциалов. Совершенно очевидно, что без знания механизмов биозлектрогенеза невозможно в полной мере оценить его роль в протекании различных жизненных функций и выработать пути использованиябиоэлектрических явлений при решении конкретных практических задач. [c.4] В связи с этим следует отметить, что Нахманзон [529, 530] создал оригинальную гипотезу "цикла ацетилхолина , объединяющую электрический и химический механизмы распространения возбуждения в нервной системе. Интересно, что ацетилхолин и холинэстераза присутствуют и в растительных тканях. Считается, что ацетилхолин у растений, как и у животных, выступает в роли регулятора биоэлектрических явлений, хотя точное его действие на эти процессы пока не известно (230]. Возможно, гипотеза единого ацетилхолинового или иного медиаторного циклического механизма возбуждения сможет найти свое подтверждение при изучении импульсной электрической активности у высших растений. По крайней мере предположение о том, что деполяризация сама способна приводить к высвобождению возбуждающей раневой субстанции при распространении ВП, выглядит, на наш взгляд, весьма правдоподобно. [c.95] Новак В.А.. Иванкина НГ. Гладкова СЕ. //Биоэлектрические явления имембранный транспорт у растений. Горький Горьк. ун-т. 1985. С. 52—57. [c.200] Я уже отмечал, что еще очень недавно исследователи были разделены на два непримиримых лагеря — сторонников сорбционной теории, возглавляемых Д. Н. Насоновым, и сторонников мембранной теории во главе с Д. Л. Рубинштейном. Темпераментные, не всегда академические споры завершились во втором поколении противников победой мембранной теории. Она восторжествовала благодаря замечательным достижениям в изучении биоэлектрических явлений нагигантских аксонах кальмаров и других удобных объектах. В самом деле, можно перфу-зировать, промыть гигантский аксон, удалить из него всю протоплазму— а способность к раздражению, к генерации нервного импульса останется. Более того,можно получить искусственную-возбудимую мембрану, обладающую всемиосновными свойствами естественной [434, 435]. Следовательно, функция аксона и. в самом деле обусловлена лишь его мембраной. Следовательно, ионная асимметрия и в самом деле создается без участия избирательной сорбции ионов в толще внутриклеточной протоплазмы. [c.100] Некоторые виды валонии, у которых большая часть таллома (редставлена пузырем, достигающим нескольких сантиметров в диа-1етре и содержащим громадную вакуоль с клеточным соком, являют-я излюбленными объектами для изучения клеточной проницаемости, ккумуляции веществ в клетке, а также биоэлектрических явлений. [c.239] Харовые водоросли представляют немалый теоретический интерес на них издавна изучали движение цитоплазмы. Кроме того, крупные клетки междоузлий этих водорослей, в которых цитоплазма ограничена постенным слоем, а центр клетки занят громадной вакуолью с клеточным соком, служат объектами для исследования биоэлектрических явлений, [c.250] В связи со сказанным выше еще раз вспомним о теории <щикла ацетилхолина , объединяющей электрический и химический механизмы распространения возбуждения в нервной системе. Как известно, ацетилхолин и холинэстераза присутствуют и в растительных тканях. Как уже отмечалось, ацетилхолин у растений и у животных выступает в роли регулятора биоэлектрических явлений, хотя для растений точное его действие на эти процессы неизвестно. [c.113] Продолжительная работа в условиях высокой внешней температурывызывает сложное течение биоэлектрических явлений сердца. В частности, после нагрузки в течение значительного времени сохраняется синусовая тахикардия, что свидетельствует о продолжаюш ейся после нагрузки напряженной деятельности сердца. Кроме того, после работы, сопровождаюш ейся перегреванием тела, наблюдаются случаи замедления внутрижелудочковой проводимости, а также может возникнуть перенапряжение сердечной мышцы и снижение функциональных возможностей сердца. [c.218]
Биоэлектрические потенциалы и токи в клетке
Измерение магнитного поля нерва. Надежное измерение магнитного поляизолированного нерва стало возможным и было осуществлено в 1980 г. Виксво с соавторами благодаря созданию специального датчика с миниатюрнойиндукционной катушкой [И, 56, 117, 166, 196, 198, 202]. При возбуждении нервной клетки и распространении вдоль аксона импульса в мембране клеткивозникают биоэлектрические генераторы, как показано на рис. 2.41, а. Для осесимметричной цилиндрической клетки эти первичные генераторы, направленные внутри мембраны радиально, порождают мембранный, внутриклеточный и внеклеточный токи и соответствующее электромагнитное полево всем рассматриваемом пространстве. Прохождение возбуждения по нерву сопровождается специфическим однофазным импульсом трансмембранного потенциала, или потенциалом действия, восходящий участок которогохарактеризует процесс деполяризации нервной клетки, а нисходящий участок — процесс ее реполяризации (в отличие от потенциала действия клеток миокарда этот импульс не имеет фазы плато между участками деполяризации и реполярнзации). Например, гигантский аксон лангуста, исследованный экспериментапьно [73, с. 78 159, с. 512], имеет потенциал действия с амплитудой около 100 мВ и длительностью около 1 мс, причем при его распространении ширина областидеполяризации в пространстве составляет около 3 мм. Поскольку скорость распространения импульса возбуждения вдоль оси клетки можно с>ш-тать постоянной, в каждый зафиксированный момент времени распределение трансмембранного потенциала вдоль осн клетки будет подобно по формеимпульсу потенциала действия во времени. При этом ток внутри аксона, направленный вдоль его оси (осевой ток), пропорционален производнойтрансмембранного потенциала по направлению оси, и его распределение имеетдвухфазную структуру. Магнитное поле клетки имеет осесимметричную форму, его линии магнитной индукции представляют собой концентрические окружности, ось которых совпа-136 [c.136] Вопрос о природе возникновения биоэлектрических потенциалов решения еще не получил. В настоящее время многие физиологи считают, что возникновение биоэлектрических потенциалов связано с неравномерным распределением ионов в тканях. Предполагают, что катионы (например, К ) легче проходят на поверхность клетки (поскольку оболочка клетки в разной мере проницаема для ионов), чем анионы. Благодаря этому создается биоэлектрический потенциал покоящейся клетки. Проницаемость клеточных мембран меняется особенно в момент возбуждения, в результате возникает ток действия. [c.169] Биоэлектрическим потенциалом называют разность потенциала, возникающего в тканях животного и растения. Опытами Гальвани (1791) было доказановозникновение электрических явлений в мышцах и нервах лягушек. На их основе впервые была высказана идея о существовании так называемого животного электричества. Исследование электрических явлений в тканях животных и растений продолжалось в XIX в. и особенно активно проводится в XX в. как в СССР, так и за рубежом. До недавнего времени считалось, что токи в организме возникают вследствие повреждения клетки поврежденный участок приобретаетотрицательный потенциал по отношению к неповрежденному. В настоящее время доказано возникновение разности потенциала и между участками неповрежденной ткани. Разность потенциалов в клетках животных организмов достигает 0,05—0,1 в. Такая же разность потенциалов обнаружена у одноклеточных животных и в растительных клетках. [c.169]
12 Биоэлектрические потенциалы
1. Ионные потоки в мембране.
2. Потенциал покоя. Уравнение Гольдмана-Ходжкина-Катца.
3. Потенциал действия и его распространение.
4. Основные понятия и формулы.
5. Задачи.
В живых организмах происходят разнообразные электрические процессы. Функционирование живых тканей сопровождается электрическими явлениями. Генерация и распространение электрических потенциалов - важнейшее физическое явление в живых клетках и тканях.
Биоэлектрический потенциал - разность потенциалов между двумя точками живой ткани, определяющая ее биоэлектрическую активность. Биопотенциал имеет мембранную природу.
12.1. Ионные потоки в мембране
Сквозь мембрану проходят потоки ионов, для которых справедливо уравнение Нернста-Планка (11.18):
![]() Эйнштейн
показал, что подвижность ионов
Um выражается
через коэффициент диффузии D и абсолютную
температуру Т: Um =
D/(RT), тогда уравнение Нернста-Планка
принимает вид
Эйнштейн
показал, что подвижность ионов
Um выражается
через коэффициент диффузии D и абсолютную
температуру Т: Um =
D/(RT), тогда уравнение Нернста-Планка
принимает вид
![]() где
с - концентрация ионов; R - универсальная
газовая постоянная.
где
с - концентрация ионов; R - универсальная
газовая постоянная.
В первом приближении можно считать, что поле внутри мембраны однородно. В этом случае dp/dx = φм/l, где L - толщина мембраны, а φм - разность потенциалов между ее поверхностями, называемая мембранным потенциалом. Теперь уравнение Нернста-Планка будет иметь вид
 Для
упрощения записи введем величину ψ,
которую называют
Для
упрощения записи введем величину ψ,
которую называют
безразмерным потенциалом:
12.2. Потенциал покоя. Стационарный потенциал Гольдмана-Ходжкина-Катца
Потенциал покоя
В нормально функционирующей клетке поддерживается наиболее благоприятный состав ионов. Различие в их концентрациях по разные стороны мембраны приводит к появлению разности потенциалов.
Потенциал покоя - разность потенциалов между цитоплазмой и окружающей средой в нормально функционирующей клетке.
В 1902 г. Бернштейн предположил, что потенциал покоя обусловлен проницаемостью мембраны для ионов К+. В состоянии покоя плотность потока ионов равна нулю, и уравнение (12.5) принимает следующий вид:
 Потенциал,
определяемый формулой (12.7),называют
равновесным потенциалом. Расчеты,
выполненные по этой формуле, существенно
расходятся с экспериментальными данными
при низких концентрациях К+. Это указывает
на то, что «калиевая» теория потенциала
несовершенна.
Потенциал,
определяемый формулой (12.7),называют
равновесным потенциалом. Расчеты,
выполненные по этой формуле, существенно
расходятся с экспериментальными данными
при низких концентрациях К+. Это указывает
на то, что «калиевая» теория потенциала
несовершенна.
Стационарный потенциал Гольдмана-Ходжкина-Катца
Причина отклонения равновесного потенциала от опытных данных заключается в проницаемости мембраны и для других ионов, которые вносят свой вклад в образование мембранного потенциала. Основной вклад в суммарный поток зарядов, а следовательно, в создание и поддержание потенциала покоя, помимо К+, вносят ионы Na+, Cl-. Суммарная плотность потока этих ионов с учетом их знаков равна
![]() Знак
«-» перед JCl- указывает
на отрицательный заряд.
Знак
«-» перед JCl- указывает
на отрицательный заряд.
В стационарном состоянии (когда параметры системы не изменяются) суммарная плотность потока равна нулю, т.е. число различных ионов, проходящих в единицу времени через мембрану внутрь клетки, равно числу ионов, выходящих из клетки через мембрану: J = 0
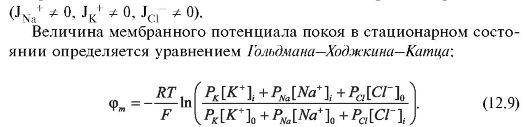 Здесь,
во избежание сложностей с индексацией,
для обозначения концентрации вместо
буквы с использованы
квадратные скобки: []i и[]0 -
концентрации соответствующих
ионов внутри и вне клетки.
Здесь,
во избежание сложностей с индексацией,
для обозначения концентрации вместо
буквы с использованы
квадратные скобки: []i и[]0 -
концентрации соответствующих
ионов внутри и вне клетки.
Очевидно, что формула для равновесного потенциала (12.7) получается из формулы стационарного потенциала (12.9) при РNa = 0 и РС1.= 0 Таким образом, уравнение Гольдмана-Ходжкина-Катца существенно уточняет теорию Бернштейна.
Большая часть сведений о нервных клетках получена при изучении аксона кальмара, достигающего почти миллиметровой толщины. Его изолированные нервные волокна довольно долго сохраняют способность передавать нервные импульсы. Рассчитаем стационарный мембранный потенциал для клеток аксона кальмара. При вычислении по формуле (12.9) вместо самих коэффициентов проницаемости можно использовать отношение между ними, которое для аксона выражается следующими числами:
|
|
 Значения
концентраций приведены ниже.
Значения
концентраций приведены ниже.
 Подставив
эти значения в формулу (12.9) при Т = 303 К
(30 °С), получим:
Подставив
эти значения в формулу (12.9) при Т = 303 К
(30 °С), получим:
 Что
достаточно хорошо согласуется со
значением, определенным опытным путем.
Что
достаточно хорошо согласуется со
значением, определенным опытным путем.