

Распространение возбуждения по безмиелиновым нервным волокнам
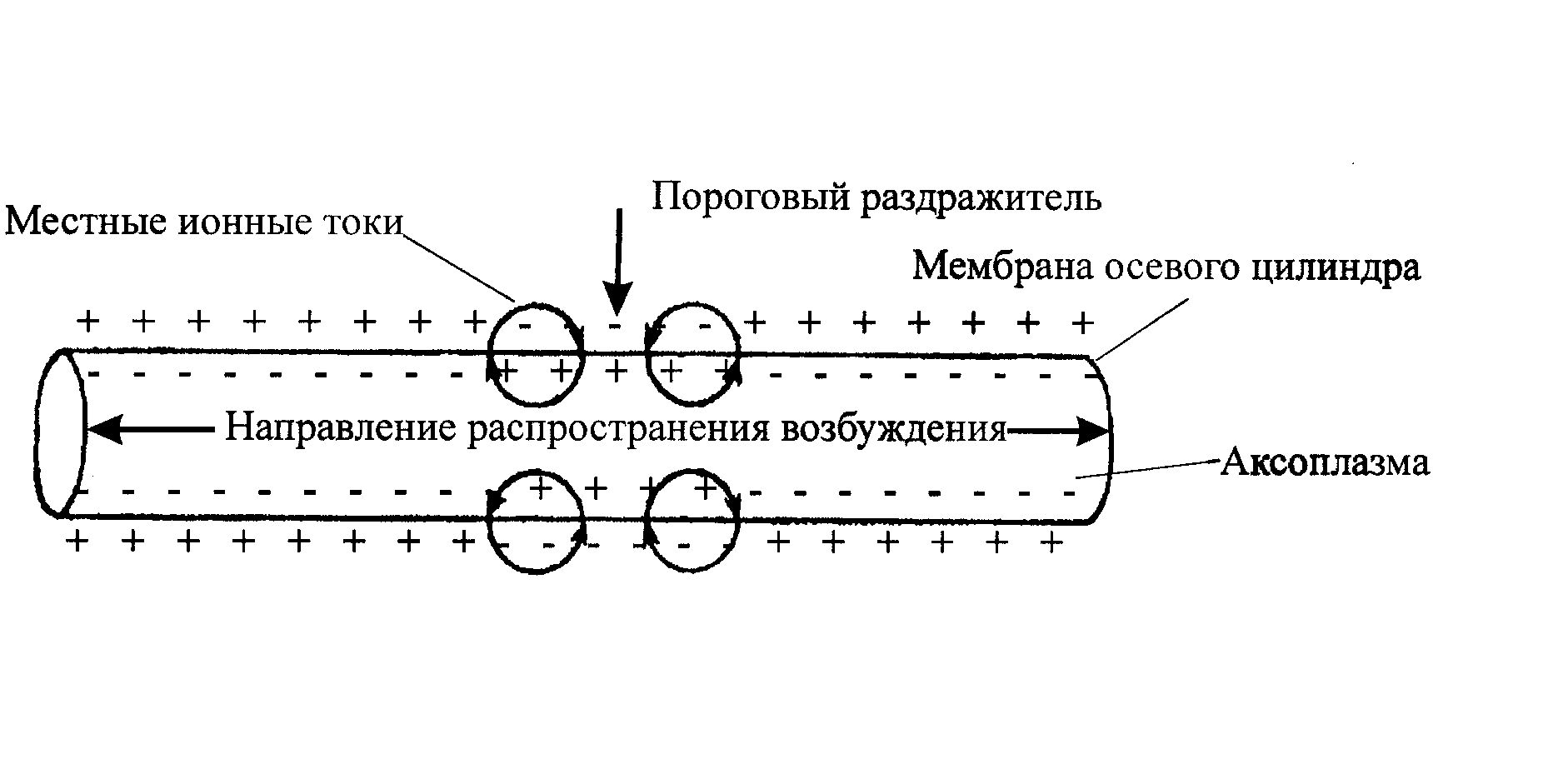
В состоянии покоя мембрана осевого цилиндра внутри заряжена отрицательно, а снаружи – положительно. При генерации ПД (во время реверсии) заряды меняются: внутри нервное волокно становится заряженным положительно, а снаружи - отрицательно. Между возбужденным и соседними невозбужденными участками мембраны осевого цилиндра возникает разность потенциалов, и начинают циркулировать местные ионные токи.
Петля местного ионного тока состоит из двух ветвей - внутренней выходящей и наружной входящей. Внутренняя ветвь проходит по аксоплазме от «+» возбужденного участка к «-» невозбужденного. Наружная ветвь направляется по межклеточной жидкости от «+» невозбужденного участка к «-» возбужденного. Выходящая из аксоплазмы ветвь местного ионного тока, обладает раздражающим действием по отношению к невозбужденному участку нервного волокна. При этом сила местного ионного тока достигает порогового или надпорогового уровня. Поэтому в невозбужденном участке генерируется ПД.
Осевой цилиндр миелинизированного нервного волокна покрыт миелиновой оболочкой, которая обладает большим электрическим сопротивлением. Миелиновая оболочка образуется специальными шванновскими клетками, которые многократно обертывают осевой цилиндр нервного волокна. Участки свободные от миелина обладают наибольшей возбудимостью и называются узловыми перехватами Ранвье.
Строение миелинизированного нервного волокна

Потенциалы действия в мякотных волокнах генерируются только в перехватах Ранвье. Поэтому возбуждение по миелинизированным нервным волокнам проводится сальтаторно (скачкообразно, лат. «saltus» – прыжок), «перепрыгивая» через участки покрытые миелином и распространяясь только по перехватам Ранвье. Такой механизм проведения возбуждения имеет два основных преимущества: большую скорость проведения ПД и малые затраты энергии.
Распространение возбуждения по миелинизированным нервным волокнам

Скорость проведения ПД зависит не только от миелинизации нервного волокна, но и от его диаметра, а также от величины фактора надежности. С увеличением диаметра сопротивление нервного волокна падает, поэтому скорость проведения импульса возрастает. При одном и том же диаметре волокна скорость проведения зависит от величины фактора надежности - отношения амплитуды ПД (мв) к критическому уровню деполяризации (мв). Чем больше показатель надежности, тем выше скорость проведения и наоборот.
По морфо-функциональным особенностям нервные волокна подразделяются на три основных типа: миелинизированные А и В, немиелинизированные С.
Волокна типа А диаметром от 1 до 22 мкм и скоростью проведения возбуждения 5-120 м/с делятся на четыре подгруппы: АА, А, А. Наибольшими скоростью проведения возбуждения и возбудимостью отличаются волокна типа А
К волокнам типа А относятся двигательные волокна, иннервирующие скелетные мышцы, афферентные волокна от проприорецепторов, терморецепторов, барорецепторов, тактильных и болевых рецепторов.
Диаметр волокон типа В 1-3,5 мкм, скорость проведения возбуждения – 3-18 м/с. К ним относятся в основном волокна вегетативной нервной системы.
Тип С составляют безмиелиновые нервные волокна диаметром 0,5-2 мкм и скоростью проведения возбуждения 0,5-3 м/с. Нервные волокна типа С характеризуются не только наименьшей скоростью проведения возбуждения, но и минимальной возбудимостью. Это преимущественно волокна вегетативной нервной системы, а также афферентные нервные волокна от терморецепторов, барорецепторов и болевых рецепторов.
Проведение возбуждения по нервным волокнам, проходящим в составе нервного ствола, подчиняется трем законам:
закону двустороннего проведения возбуждения,
закону изолированного проведения возбуждения,
закону физиологической и анатомической целостности нервного волокна.
Согласно закону двустороннего проведения, возбуждение по нервному волокну от места нанесения раздражения распространяется в двух направлениях - как центробежно (от нервного центра), так и центростремительно (к нервному центру). Закон двустороннего проведения является искусственным экспериментальным феноменом, т.к. в рефлекторных дугах благодаря наличию синапсов возбуждение передается только в одном направлении.
Согласно закону изолированного проведения, возбуждение по нервным волокнам в составе нервного ствола проводится изолированно и с одного волокна на другие в пределах общего нервного ствола не распространяется. Это связано с тем, что высокое сопротивление нервных волокон и шунтирующие (рассеивающие) свойства межклеточной жидкости не позволяют местным ионным токам возбуждать соседние нервные волокна.
Согласно закону анатомической и физиологической непрерывности нерва, обязательным условием проведения возбуждения по нервному волокну является анатомическая и функциональная целостность мембраны осевого цилиндра. Возбуждение по нервному волокну проводится только в том случае, если оно не повреждено и сохранены все его физиологические свойства.
В участке нерва, подвергшегося повреждению (альтерации), возникает парабиоз (греч. «para» - около, «bios» - жизнь) - местное, стойкое, не колеблющееся возбуждение, которое появляется в результате воздействия альтерирующего фактора (анестетиков, электрического тока, гипоксии, воспаления, охлаждения и др.) и приводит к изменению физиологических свойств возбудимой ткани.
Российским физиологом Н.Е. Введенским (1901) в опытах на нервно-мышечном препарате выявлено три последовательных фазы парабиоза: уравнительная – парадоксальная - тормозная.
Импульсы, которые генерируются в парабиотическом участке при раздражении нерва, характеризуются удлинением всех фаз ПД, а значит, увеличением продолжительность циклов восстановления возбудимости. Чем продолжительнее и сильнее действие альтерирующего фактора, тем больше во времени растягивается ПД и тем существеннее изменения физиологических свойств ткани.
Если раздражать нерв интактного нервно-мышечного препарата импульсным электрическим током, то амплитуда мышечного сокращения будет прямо пропорциональна частоте стимуляции: при стимуляции высокой частотой амплитуда мышечного сокращения будет больше, чем при раздражении низкой.