

Environmental Biotechnology - Theory and Application - G. M. Evans & J. C. Furlong
.pdf54 Environmental Biotechnology
A number of species, for example, accumulate potassium chloride (KCl) in their cytoplasm, with the concomitant result that extremozymes isolated from these organisms will only function properly in the presence of high KCl levels. By the same token, many surface structural proteins in halophiles require severely elevated concentrations of sodium salts.
Acidophiles thrive in the conditions of low pH, typically below 5, which occur naturally as a result of sulphurous gas production in hydrothermal vents and may also exist in residual spoils from coal-mining activity. Though they can tolerate an externally low pH, an acidic intra-cellular environment is intolerable to acidophilic organisms, which rely on protective molecules in, or on, their cell walls, membranes or outer cell coatings to exclude acids. Extremozymes capable of functioning below pH1 have been isolated from these structures in some acidophile species.
At the other end of the spectrum, alkaliphiles are naturally occurring species which flourish in soda lakes and heavily alkaline soils, typically enduring pH9 or more. Like the previous acidophiles, alkaliphiles require more typically neutral internal conditions, again relying on protective chemicals on or near their surfaces or in their secretions to ensure the external environment is held at bay.
Diverse degradative abilities
Bacteria possessing pathways involved in the degradation of a number of organic molecules of industrial importance, have been acknowledged for some time. One oft-quoted example is that for toluene degradation in Pseudomonas putida, which exhibits a fascinating interplay between the genes carried on the chromosome and the plasmids (Burlage, Hooper and Sayler 1989). Bacteria are constantly being discovered which exhibit pathways involved in the degradation and synthesis of chemicals of particular interest to environmental biotechnologists. For example, a new class of biopolymer produced by the bacterium, Ralstonia eutropha, containing sulphur in its backbone, has recently been identified. (Lutke¨-Eversloh et al. 2001) It is possible that these and other novel biopolymers awaiting discovery, will have innovative and exciting applications in clean technology.
In very recent years, bacteria representing very diverse degradative abilities have been discovered in a variety of niches adding almost daily, to the pool of organisms of potential use to environmental biotechnology. By illustration these include phenol-degrading Oceanomas baumannii isolated from estuarine mud from the mouth of the River Wear, UK (Brown, Sutcliffe and Cummings 2001), chloromethane utilising Hyphomicrobium and Methylobacterium from polluted soil near a petrochemical factory in Russia (McDonald et al. 2001) and a strain of Clostridium able to degrade cellulose, isolated from soil under wood chips or the forest floor in northeast USA. In addition to their cellulytic activity, these Clostridia were also found to be mesophilic, nitrogen-fixing, spore-forming and obligate anaerobes (Monserrate, Leschine and Canale-Parola 2001). Again, there is interest in this organism with regard to clean technology in the hope that it may
Fundamentals of Biological Intervention 55
be used to convert cellulose into industrially useful substances. A note of caution is that cellulose is a major product of photosynthesis and, being the most abundant biopolymer on this planet, has a vital role to play in the carbon cycle. Large-scale disturbance of this balance may have consequences to the environment even less welcome than the technologies they seek to replace. However, judicious use of this biotechnology could reap rewards at many levels.
Bacteria have also adapted to degrade man-made organics called xenobiotics.
Xenobiotics and Other Problematic Chemicals
The word is derived from the Greek ‘xenos’ meaning foreign. Throughout this book the definition used is that xenobiotics are compounds which are not produced by a biological procedure and for which no equivalent exists in nature. They present a particular hazard if they are subject to bioaccumulation especially so if they are fat soluble since that enables them to be stored in the body fat of organisms providing an obvious route into the food chain. Despite the fact that these chemicals are man made, they may still be degraded by micro-organisms if they fit into one of the following regimes; gratuitous degradation, a process whereby the xenobiot resembles a natural compound sufficiently closely that it is recognised by the organism’s enzymes and may be used as a food source, or cometabolism where the xenobiot is degraded again by virtue of being recognised by the organism’s enzymes but in this case its catabolism does not provide energy and so cannot be the sole carbon source. Consequently, cometabolism may be sustained only if a carbon source is supplied to the organism. The ability of a single compound to be degraded can be affected by the presence of other contaminants. For example, heavy metals can affect the ability of organisms to grow, the most susceptible being Gram positive bacteria, then Gram negative. Fungi are the most resistant and actinomycetes are somewhere in the middle. This being the case, model studies predicting the rate of contaminant degradation may be skewed in the field where the composition of the contamination may invalidate the study in that application. Soil micro-organisms in particular are very versatile and may quickly adapt to a new food source by virtue of the transmission of catabolic plasmids. Of all soil bacteria, Pseudomonads seem to have the most highly developed ability to adapt quickly to new carbon sources. In bacteria, the genes coding for degradative enzymes are often arranged in clusters, or operons, which usually are carried on a plasmid. This leads to very fast transfer from one bacterium to another especially in the case of Pseudomonas where many of the plasmids are self-transmissible. The speed of adaptation is due in part to the exchange of plasmids but in the case of the archaeans particularly, the pathways they carry, which may have been latent over thousands of bacterial generations, owe their existence to previous exposure over millions of years to an accumulated vast range of organic molecules. It is suggested that, unless there has been evolutionary pressure to the contrary, these latent pathways are retained to

56 Environmental Biotechnology
a large extent requiring little modification if any to utilise new xenobiotics. Even so, bioremediation may require that organisms are altered in some way to make them more suitable for the task and this topic is addressed in Chapter 9. Briefly, the pathways may be expanded by adaptation to the new molecule, or very much less commonly, wholescale insertion of ‘foreign’ genes may occur by genetic manipulation. There have been several cases reported where catabolic pathways have been expanded in the laboratory. Hedlund and Staley (2001) isolated a strain of Vibrio cyclotrophicus from marine sediments contaminated with creosote. By supplying the bacteria with only phenanthrene as a carbon and energy source, the bacteria were trained to degrade several PAHs although some of these only by cometabolism with a supplied carbon source.
Endocrine disrupters
To date, there are chemicals, including xenobiotics, which still resist degradation in the environment. This may be due to a dearth, at the site of contamination, of organisms able to degrade them fully or worse, microbial activity which changes them in such a way that they pose a bigger problem than they did previously. One such example is taken from synthetic oestrogens such as 17α-ethinyloestradiol commonly forming the active ingredient of the birth control pills, and the natural oestrogens which, of course, are not xenobiotics. Natural oestrogens are deactivated in humans by glucuronidation, as shown in Figure 3.1, which is a conjugation of the hormone with UDP-glucuronate making the compound more
Figure 3.1 Glucuronidation
Fundamentals of Biological Intervention 57
polar and easily cleared from the blood by the kidneys. It is in this modified and inactive form that it is excreted into the sewage. However, bacteria present in the aerobic secondary treatment in sewage treatment plants, have the enzyme,
β-glucuronidase, which removes this modification thus reactivating the hormone. As an aside, glucuronidation is not confined to hormones but is a process used
to detoxify a number of drugs, toxins and carcinogens in the liver. The enzyme catalysing this process is induced in response to prolonged exposure to the toxin thus imparting increased tolerance or even resistance to the chemical.
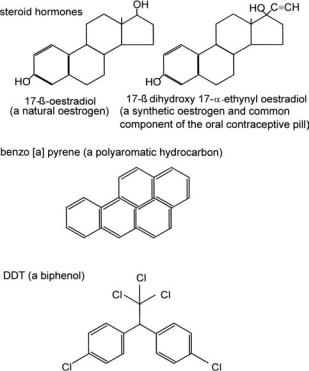
Returning to the problem of elevated levels of active hormones in the waterways, another aspect is that steroids do not occur in bacteria, although they are present in fungi, and so bacteria lack the necessary pathways to allow complete degradation of these hormones at a rate compatible with the dwell time in sewage treatment plants. The consequence has been raised levels of reactivated oestrogen and 17α-ethinyloestradiol in the waterways leading to disturbances of the endocrine, or hormonal, system in fauna downstream from sewage treatment plants. Such disturbances have been monitored by measuring the presence of the protein vitellogenin (Sole et al. 2001) which is a precursor to egg yolk protein, the results of which have indicated feminisation of male fish in many species including minnows, trout and flounders. The source of environmental oestrogens is not confined to outfall from sewage treatment plants, however, the fate of endocrine disrupters, examples of which are given in Figure 3.2, in sewage treatment plants is the subject of much research (Byrns 2001). Many other chemicals, including polyaromatic hydrocarbons (PAHs), dichlorodiphenyltrichloroethane (DDT), alkyl phenols and some detergents may also mimic the activity of oestrogen. There is general concern as to the ability of some organisms to accumulate these endocrine disrupters in addition to the alarm being raised as to the accumulative effects on humans of oestrogen-like activity from a number of xenobiotic sources.
To date there is no absolute evidence of risk to human health but the Environmental Agency and Water UK are recommending the monitoring of environmental oestrogens in sewage treatment outfall. Assays are being developed further to make these assessments (Gutendorf and Westendorf 2001) and to predict potential endocrine disrupter activity of suspected compounds (Takeyoshi et al 2002). Oestrogen and progesterone are both heat labile. In addition, oestrogen appears to be susceptible to treatment with ultra-violet light, the effects of which are augmented by titanium dioxide (Eggins 1999). The oestrogen is degraded completely to carbon dioxide and water thus presenting a plausible method for water polishing prior to consumption.
Another method for the removal of oestrogens from water, in this case involving Aspergillus, has also been proposed (Ridgeway and Wiseman 1998). Sulphation of the molecule by isolated mammalian enzymes, as a means of hormone inactivation is also being investigated (Suiko 2000). Taken overall, it seems unlikely that elevated levels of oestrogen in the waterways will pose a problem
58 Environmental Biotechnology
Figure 3.2 Endocrine disrupters
to human health in drinking water although, this does not address the problem affecting hormone-susceptible organisms living in contaminated water and thus exposed to this potential hazard.
New discoveries
Almost daily, there are novel bacteria being reported in the literature which have been shown to have the capacity to degrade certain xenobiots. Presumably the mutation which occurred during the evolution of the organism conferred an advantage, and selective pressure maintained that mutation in the DNA, thus producing a novel strain with an altered phenotype. Some example of such isolates are described here. Reference has already been made to some PAHs mimicking oestrogen which earns those chemicals the title of ‘endocrine disrupters’. This is in addition to some being toxic for other reasons and some being carcinogenic or teratogenic. The PAHs are derived primarily from the petrochemicals industry and are polycyclic hydrocarbons of three or more rings which include as members, naphthalene and phenanthrene and historically have been associated
Fundamentals of Biological Intervention 59
with offshore drilling, along with alkylphenols. Several genera of bacteria are now known to be able to degrade PAHs and recently, a novel strain of Vibrio cyclotrophicus able to digest naphthalene and phenanthrene, was isolated from creosote-contaminated marine sediments from Eagle Harbour, Washington, USA. It would appear that bacteria isolated from the same marine or estuarine environments may vary quite considerably in their abilities to degrade certain PAHs. This observation is viewed as indicative of diverse catabolic pathways demonstrated by these organisms and awaiting our full understanding (Hedlund and Staley 2001).
Polycyclic hydrocarbons (PCBs) are xenobiotics which, due to their high level of halogenation, are substrates for very few pathways normally occurring in nature. However, a strain of Pseudomonas putida able to degrade PCBs, was isolated recently from wastewater outflow from a refinery. This was achieved by the bacterium employing two pathways encoded by two separate operons; the tod pathway employed in toluene degradation, and the cmt pathway which normally is responsible for the catabolism of p-cumate which is a substituted toluene. The mutation which allowed this strain to utilise the cmt pathway was found to be a single base change to the promoter-operator sequence. This allowed all the enzymes in this pathway to be expressed under conditions where their synthesis would normally be repressed. Thus, the two pathways could work in conjunction with each other to metabolise PCBs, a relationship described as mosaic (Ohta et al 2001).
The pthalates are substituted single-ring phenols and include terephthalic acid and its isomers, the major chemicals used in manufacture of polyester fibres, films, adhesives, coatings and plastic bottles. In Chapter 2, and earlier in this chapter, homage has been paid to the resources of genetic capability exhibited by the archaeans. In a recent analysis of anaerobic sewage sludge, a methanogenic consortium of over 100 bacterial clones were found to have the capability to digest terephthalate. Characterisation of these by analysis of their ribosomal DNA sequences, revealed that almost 70% were archaeans most of which had not been previously identified, and that nearly 90% of the total bacteria comprised two of the novel archaean species. These two species are believed to be responsible for the degradation of terephthalic acid (Wu et al 2001). During wastewater treatment, terepthalic acid is usually treated by aerobic processes. However, this consortium, or others like it provide an anaerobic alternative which, being methanogenic, may be structured to offset processing costs by the utilisation of the methane.
Mobility of DNA
Throughout this book, reference is made to the movement of genes within and between organisms. The reason why it appears at all in a book on environmental biotechnology is to emphasise the ‘oneness’ of the environment, not just at the more obvious level of industrial impact but right down to the interaction
60 Environmental Biotechnology
between the genetic material of organisms themselves. Plasmids may be transferred between bacteria by conjugation, of which there are several types, but all of which require direct cell to cell contact. Not only are genes transferred between bacteria on plasmids, but bacteriophages (bacterial viruses) are also vectors for intercellular transmission. Similarly, eukaryotic viruses are able to transfer genetic material between susceptible cells. In addition, bacterial cells may pick up DNA free in the environment under conditions where their cell wall has become ‘leaky’ to fragments of this macromolecule, a process called transformation. There is also considerable rearrangement of genomic material within an organism stimulated by the presence of transposons. There are many classes of transposable elements which are short pieces of DNA, able to excise themselves, or be excised, out of a genome. Often they take with them neighbouring pieces of DNA, and then reinsert themselves, sometimes with the assistance of other genes, into a second site distinct from the original location on the same genome. Insertion may be into specific sites or random, depending on the nature of the transposon. Transposition normally requires replication of the original DNA fragment and so a copy of this transposon is transferred leaving the original behind. Transposition is widespread and occurs in virtually all organisms for which evidence of this process has been sought, both prokaryotic and eukaryotic. The term ‘transposable element’, was first coined by Barbara MacClintock, who discovered them in maize, publishing her data in the early 1950s. However, it was not until many years later that the full significance of her work was being recognised, with similar elements being discovered in bacteria. Transposable elements are known to promote the fusion of plasmids within a bacterial cell, where more than one type of plasmid is present. The fusion is stimulated by the presence of insertion sequences (IS), which are short pieces of DNA of a defined and limited range of sequences. They are often found at either end of a transposable element. Their presence enables various DNA rearrangements to take place leading to moderation of gene expression. Taking together the reorganisation of DNA within all types of organisms attributable to transposable elements and IS, with transfer of DNA between organisms by plasmids and transformation, in the case of prokaryotes, and viruses in the case of both prokaryotes and eukaryotes, the potential for DNA rearrangement within and between organisms is enormous.
It has been proposed (Reanney 1976), that such transfer is far more universal than had previously been voiced. Transfer of genes by extra chromosomal elements (ECEs), which is the all-embracing name given to include plasmids and viruses, models the means by which molecular evolution takes place in the environment. The proposal is that the evolutionary process occurs principally by insertions and deletions of the genome such as those caused by the activities of ECEs and transposable elements and not by point mutations more frequently observed in isolated cultures such as those maintained in laboratory conditions. It is further suggested that much of the phenotypic novelty seen in evolution is the result of rearrangement of existing structural genes into a different region
Fundamentals of Biological Intervention 61
of the genome and therefore operating under different parameters affecting gene regulation. Transfer of genes across wide taxonomic gaps is made possible by the mobile nature of ECEs many of which may cross species barriers often resulting in the insertion of all or part of the ECE into the recipient genome. Examples of such mobility are viruses which infect a wide host range, such as some retroviruses, the alfalfa mosaic virus, and the Ti plasmid of Agrobacterium tumefaciens which the bacterium introduces into plant cells. The retroviruses, of which Human Immunodeficiency Virus (HIV) is an example, are unusual in having RNA as their genetic material. They replicate in a manner which includes double-stranded DNA as an intermediate and so may integrate into the host cell genome. RNA viruses tend to be more susceptible than DNA viruses to mutation presumably due to the less chemically stable nature of the macromolecule. They have been invoked by Reanney (1976) as being the likely agents for the spread of genetic information between unrelated eukaryotes. His observations led him to conclude that there is only a blurred distinction between cellular and ECE DNA both in eukaryotes and prokaryotes and further suggest that no organism lives in true genetic isolation as long as it is susceptible to at least one of the classes of ECEs described above. Clearly, for the mutation to be stabilised, it must occur in inheritable DNA sequences, a situation reasonably easy to achieve in microbes and at least possible in multicellular organisms.
The existence of genetic mobility has been accepted for many years, even though the extent and the mechanisms by which it operates are still being elucidated. From this knowledge several lessons may be learned; among them, that the genetic environment of any organism may well be significant and that there is some justification in viewing the principle of genetic engineering as performing in the laboratory, a process which is occurring in abundance throughout the living world. This topic is explored further in Chapters 9, 10 and 11.
Closing Remarks
As has been seen, even within the brief discussion in this chapter, life on Earth is a richly varied resource and the functional reality of biodiversity is that many more metabolic pathways exist, particularly within the microbial melting pot, than might be commonly supposed. As a result, a number of generally unfamiliar groups of chemicals and organisms have implications for the application of environmental biotechnology which exceed their most obvious contributions to a wider consideration of the life sciences. Hence, xenobiotics, as an example of the former, represent a current problem for which the solution remains unresolved and extremophiles, as the latter, hold the potential to revolutionise many industrial procedures, thereby heralding major benefits in terms of ‘clean technology’. There are many aspects of current environmental management for which there is no presently relevant biotechnological intervention. However, this is not a static science, either in terms of what can be done, or the tools available. Discoveries
62 Environmental Biotechnology
and developments both within the field and from other disciplines can and do filter in and alter the state of the possible. Thus, while the bulk of the rest of this book addresses the sorts of biotechnological applications routinely applied to deal with today’s environmental problems, it must be borne in mind that some of the issues described in this section may well assume greater relevance in the future and this is a point which will, at least in part, be reviewed in the final chapter.
References
Brown, G.R., Sutcliffe, I.C. and Cummings, S.P. (2001) Reclassification of [Pseudomonas] doudoroffi (Baumann et al. 1983) into the genus Oceanomonas gen. nov. as Oceanomonas doudoroffi comb. nov., and description of a phenoldegrading bacterium from estuarine water as Oceanomonas baumannii sp. nov., International Journal of Systemic and Evolutionary Microbiology, 51: 67 – 72.
Burlage, R.S., Hooper, S.W., and Sayler, G.S. (1989) The Tol (PWWO) catabolic plasmid, Applied and Environmental Microbiology, 55: 1323 – 8.
Byrns, G. (2001) The fate of xenobiotic organic compounds in wastewater treatment plants, Water Research, 35: 2523 – 33.
Eggins, B. (1999), EDIE news, www.edie.net/index2.html.
Gutendorf, B. and Westendorf, J. (2001) Comparison of an array of in vitro assays for the assessment of the estrogenic potential of natural and synthetic estrogens, phytoestrogens and xenoestrogens, Toxicology, 166: 79 – 89.
Hedlund, B.P. and Staley, J.T. (2001) Vibrio cyclotrophicus sp. nov., a polycyclic aromatic hydrocarbon (PAH) – degrading marine bacterium, International Journal of Systemic and Evolutionary Microbiology, 51: 61 – 6.
Lutke¨-Eversloh, T., Bergander, K., Luftmann, H. and Steinbuche,¨ H. (2001) Identification of a new class of biopolymer: bacterial synthesis of a sulphurcontaining polymer with thioester linkages, Microbiology, 147: 11 – 19.
McDonald, I.R., Doronina, N.V., Trotsenko, Y.A., McAnulla, C. and Murrell, J.C. (2001) Hyphomicrobium chloromethanicum sp. nov. and Methylobacterium chloromethanicum sp. nov., chloromethane-utilizing bacteria isolated from a polluted environment International Journal of Systemic and Evolutionary Microbiology, 51: 119 – 22.
Monserrate, E., Leschine, S.B. and Canale-Parola, E. (2001) Clostridium hungatei sp. nov., a mesophilic, N2-fixing cellulolytic bacterium isolated from soil, International Journal of Systematic and Evolutionary Microbiology, 51: 123 – 32.
Ohta, Y., Maeda, M. and Kudo, T. (2001) Microbiology, 147: 31 – 41.
Reanney, D. (1976) Extrachromosomal elements as possible agents of adaptation and development, Bacteriological Reviews, 40: 552 – 90.
Ridgeway, T.J. and Wiseman, H. (Nov 1998) Biochemical Society Transactions, 26(4): 675 – 80.
Fundamentals of Biological Intervention 63
Sole, M., Porte, C. and Barcelo, D. (2001) analysis of the estrogenic activity of sewage treatment works and receiving waters using vitellogenin induction in fish as a biomarker, TRAC Trends in Analytical Chemistry, 20: 518 – 25.
Suiko, M., Sakakibara, Y. and Liu, M.C. (2000) Sulfation of environmental estrogen-like chemicals by human cytosolic sulfotransferases, Biochemical and Biophysical Research Communications, 267(1): 80 – 4.
Takeyoshi, M., Yamasaki, K., Sawaki, M., Nakai, M., Noda, S. and Takatsuki, M. (2002) The efficacy of endocrine disrupter screening tests in detecting anti-estrogenic receptor-ligand effects downstream of interactions, Toxicology Letters, 126: 91 – 8.
Wright, P.C. and Raper, J.A. (Feb. 1996) A review of some parameters involving fluidized bed bioreactors, Chemical Engineering Technology, 19: 50 – 64.
Wu, J.H., Liu, W.T., Tseng, I.C. and Cheng, S.S. (2001) Characterization of microbial consortia in a terephthalate-degrading anaerobic granular sludge system, Microbiology, 147: 373 – 82.
Case Study 3.1 Feminisation of Fish (England and Wales)
Of all the endocrine disrupters, perhaps most concern has been voiced over the possible consequences of synthetic oestrogens. The most recent study into hormone disruption of fish in the UK, published by the Environment Agency (EA), has revealed that oestrogenic steroids in sewage effluent are more potent in this respect than had been previously thought. As result of this five-year investigation of 10 river catchments in England and Wales which receive sewage treatment effluent, it has become clear that the reproductive health of male fish is being adversely affected. With nearly half of all those studied either having eggs in their testes or developed female reproductive ducts, 25% producing damaged sperm, and 10% being sterile, the future sustainability of fish populations in a number of rivers in England and Wales could be at risk.
Many oestrogens, both natural and manufactured, have been implicated, but the synthetic steroid 17α ethinyloestradiol, which is present in the urine of women using the contraceptive pill, is of particular concern, producing effects in fish at concentrations below 1 µg/l.
While the resulting level of concern over this ‘feminisation’ is likely to drive the development of a risk management strategy for oestrogens in sewage effluent, which may ultimately necessitate changes in treatment practice or technology, it is clear that not all fish populations are exposed to the same degree and so careful targeting of any intended action will be essential.
The research, which was undertaken by scientists from Brunel and Exeter Universites, and funded by the EA, the Department for Environment, Food and Rural Affairs (DEFRA) and the Natural Environment Research Council (NERC), focused on wild populations of roach (Rutilus rutilus) and gudgeon (Gobio gobio). It specifically examined the causes and consequences of feminisation in these species and, while it is expected that the situation is similar for other fish species, it was not designed to draw conclusions regarding likely human health effects. In the UK, around a third of
Continued on page 64