-
Гладкие мышцы
Гладкие мышцы находятся в стенках внутренних органов, кровеносных
и лимфатических сосудов, радужной оболочки глаза, ресничном теле. Ониобразованы веретенообразными клетками, между которыми имеется осо-
бый вид плотных контактов — нексусы. Нексусы делают возможным элек-
тротоническое распространение возбуждения по мышце.
МПП гладких мышц — 60—70 мВ. Некоторые клетки имеют неус-
тойчивый МПП, который спонтанно понижается, и при достижении
КУД происходит генерация ПД. Такие клетки называются пейсмекера-
ми. Они формируют очаги автоматии в гладких мышцах. Возбуждение
электротонически распространяется от клетки к клетке со скоростью
2—10 см/с.
Развитие фазы деполяризации ПД связано с входом в клетку ионов
кальция через медленные кальций-натриевые каналы. ПД имеет боль-
шую длительность — от 50 до –250 мс.
Гладкая мышца не имеет поперечной исчерченности, т. к. миофибрил-
лы расположены неупорядоченно. Миофибриллы состоят из сократитель-
ных белков актина и миозина. Сокращение миофибрилл происходит в ре-
зультате скольжения нитей актина вдоль нитей миозина. Из регуляторных
белков присутствует только тропомиозин. Плохо развит саркоплазматиче-
ский ретикулум, поэтому кальций, необходимый для электромеханическо-
го сопряжения, поступает из внеклеточной жидкости во время ПД. Ско-
рость сокращения низкая, и при частоте 1 имп/с возникает гладкий тетанус.
Энерготраты крайне невелики, поэтому гладкие мышцы приспособлены
к длительному сокращению без развития утомления. Характерным свойст-
вом гладких мышц является пластичность, т. е. они могут быть полностью
расслаблены как в укороченном, так и в растянутом состоянии.
Регуляция тонуса и сократительной активности гладких мышц осу-
ществляется вегетативной нервной системой (ВНС), а также гумораль-
ными и механическими факторами. Гладкие мышцы обладают высокой
чувствительностью к биологически активным веществам крови и тка-
невой жидкости, т. к. имеют рецепторы на всей поверхности мембраны,
а не только в области синаптических контактов.
Некоторые особенности нервных влияний на гладкие мышцы связа-
ны со строением периферических вегетативных синапсов. Аксоны пост-
ганглионарных нейронов, направляясь к гладкой мышце, образуют сеть
варикозных расширений. Расширения заполнены синаптическими пу-
зырьками и могут формировать контакты с большим количеством глад-
комышечных клеток. Гладкомышечные клетки, расположенные далеко
от хода нервных волокон, возбуждаются потенциалами действия сосед-
них миоцитов, которые электротонически распространяются по мем-
бране и передаются через нексусы.
-
Нервная система
НЕРВНАЯ СИСТЕМА, сложная сеть структур, пронизывающая весь организм и обеспечивающая саморегуляцию его жизнедеятельности благодаря способность реагировать на внешние и внутренние воздействия (стимулы). Основные функции нервной системы – получение, хранение и переработка информации из внешней и внутренней среды, регуляция и координация деятельности всех органов и органных систем. У человека, как и у всех млекопитающих, нервная система включает три основных компонента: 1) нервные клетки (нейроны); 2) связанные с ними клетки глии, в частности клетки нейроглии, а также клетки, образующие неврилемму; 3) соединительная ткань Рефлекс — это ответная реакция организма на воздействие внешних
или внутренних раздражителей, вызывающих активацию рецепторов,
которая осуществляется ЦНС и проявляется в возникновении, измене-
нии или прекращении активности органов и тканей. Рефлекс контроли-
руется механизмом обратной связи.
Морфологической основой рефлекса является рефлекторная дуга,
которая представляет собой цепочку нейронов, связанных синаптиче-
скими контактами.
Компоненты рефлекторной дуги:
рецепторы, афферентный путь,
центральное звено,
эфферентный путь,
эффекторный орган.
Рецепторы — это специализированные образования, способные
воспринимать сигналы внешней или внутренней среды и преобразовы-
вать энергию раздражителя в электрический процесс.
Классификация рецепторов
1. По локализации раздражителя: а) экстерорецепторы — раздражи-
тель действует из внешней среды; б) интерорецепторы — воспринима-
ют изменения внутренней среды; в) проприорецепторы — рецепторы
скелетных мышц, сухожилий и суставов.
2. По виду стимулов: а) механорецепторы, б) хеморецепторы,
в) фоторецепторы.
3. По характеру ощущений: а) зрительные, б) слуховые,
в) обонятельные, г) вкусовые, д) осязательные (тактильные),
е) терморецепторы, ж) болевые (ноцирецепторы), з) рецепторы положе-
ния тела в пространстве (проприорецепторы и вестибулорецепторы).
4. По способности адаптироваться к действию раздражителя:
а) быстроадаптирующиеся (тактильные, зрительные),
б) медленноадаптирующиеся (проприорецепторы).
5. По порогу раздражения: а) низкопороговые, б) высокопороговые.
6. По характеру контакта со средой: а) дистантные (фоторецепторы,
слуховые, обонятельные, б) контактные (вкусовые, тактильные).
7. По строению рецептора: а) первичночувствующие — функцию
рецептора выполняет окончание дендрита чувствительного нейрона;
могут быть свободными или инкапсулированными,
б) вторичночувствующие — рецепторную функцию выполняет специа-
лизированная клетка, как правило, эпителиального происхождения, ко-
торая синаптически связана с дендритом сенсорного нейрона. Это зри-
тельные, слуховые, вкусовые и вестибулярные рецепторы.
Механизм возбуждения первичночувствующих рецепторов
При действии стимула на рецепторную мембрану происходит по-
вышение проницаемости для ионов натрия или кальция, ионы поступа-
ют внутрь нервного окончания, мембрана деполяризуется и формирует-
ся рецепторный потенциал. Рецепторный потенциал обладает всеми свойствами местного возбуждения (зависит от силы раздражителя, спо-
собен к суммации, распространяется с затуханием). Затем рецепторный
потенциал с помощью местных токов вызывает генерацию ПД в аффе-
рентном волокне. Частота ПД зависит от амплитуды рецепторного по-
тенциала.
Механзм возбуждения 2 чувствующих рецепторов
В рецепторной клетке под действием раздражителя открываются на-
триевые или кальциевые каналы, что приводит к возникновению рецеп-
торного потенциала. Возбуждение клетки вызывает секрецию медиато-
ра (ацетилхолин, глутаминовая кислота), и на мембране чувствительно-
го нейрона формируется генераторный потенциал. Генераторный
потенциал с помощью местных токов действует на мембрану аффе-
рентного волокна, где и возникает ПД.
Афферентное звено рефлекторной дуги представлено сенсорными
нейронами, тела которых находятся в чувствительных ганглиях спин-
номозговых нервов или соответствующих черепномозговых нервах.
Центральное звено рефлекса представлено интернейронами, кото-
рые образуют малые возбуждающие и тормозные нейронные сети
(см. выше).
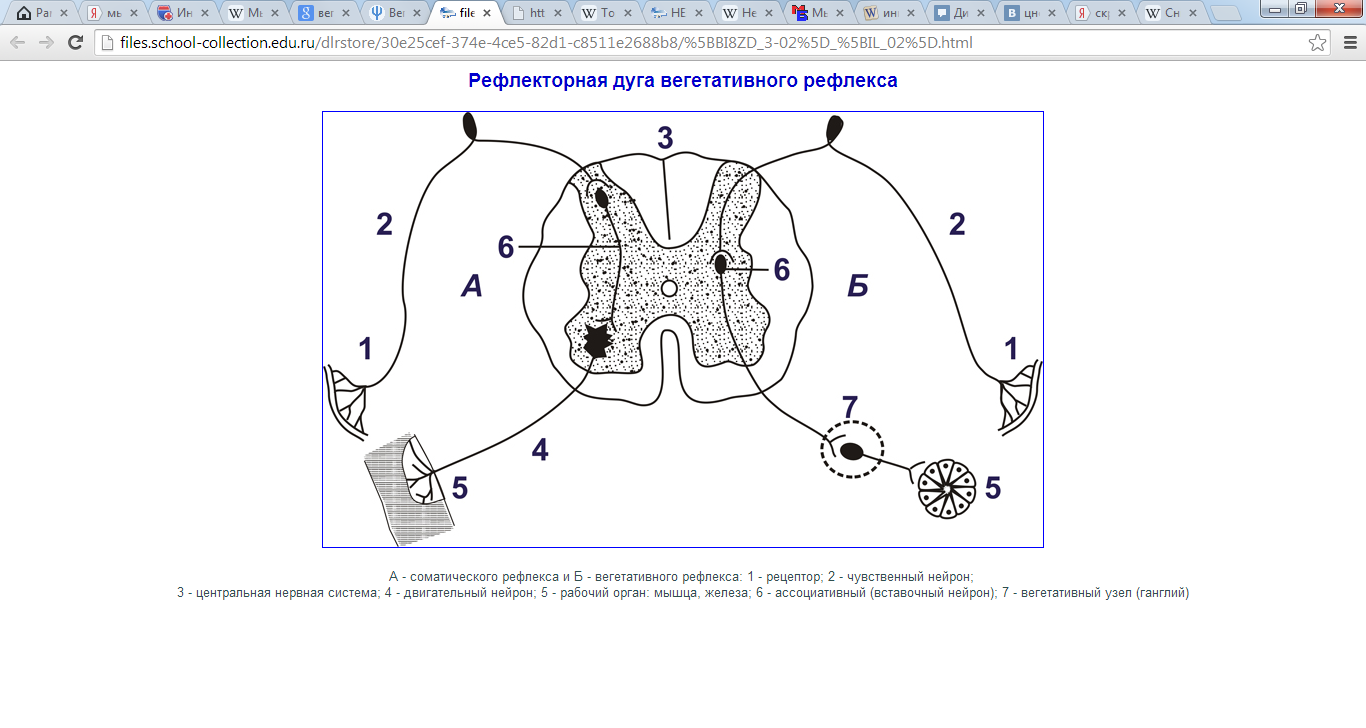
Эфферентное звено представлено одним (для соматических рефлек-
сов) или двумя (для вегетативных рефлексов) нейронами.
Эффекторный орган в соматических рефлексах — это скелетные
мышцы, в вегетативных — гладкие мышцы, железы, кардиомиоциты.
-
Нервный центр — это совокупность нейронов разных уровней ЦНС,
деятельность которых осуществляет регуляцию функций. Это функ-
циональное, а не анатомическое образование, и в основе его деятельно-
сти лежат возбуждающие и тормозные нейронные сети. Принципы
функционирования нервных сетей одинаковы во всех нервных центрах.
Свойства
1. Центральная задержка — время с момента поступления информа-
ции в нервный центр до выхода информации из нервного центра. Обу-
словлено наличием синаптической передачи в нервном центре и обра-
боткой информации.
2. Одностороннее проведение информации в нервной сети — в каж-
дом нервном центре имеются входные нейроны и нейроны на выходе,
которые не могут изменить свою функциональную принадлежность, так
как связаны синаптическими контактами.
3. Дивергенция — расхождение информации от одного нейрона
к нескольким, так как аксон одного нейрона имеет множество колате-
ралей (разветвлений), которые заканчиваются синапсами на других
нейронах.
4. Иррадиация возбуждения — следствие дивергенции, при раздраже-
нии одного афферентного входа возникает генерализованная реакция за
счет возбуждения большого количества вставочных и моторных нейро- нов. Имеет большое значение при формировании ответных реакций ор-
ганизма на действие сильных и биологически значимых раздражителей.
5. Конвергенция — схождение информации от разных нейронов
к одному интернейрону или мотонейрону. Если информация от многих
нейронов поступает на моторные нейроны, то этот частный случай кон-
вергенции носит название общий конечный путь или воронка Шер-
рингтона.
6. Суммация возбуждения в нервном центре является проявлением
интегративной деятельности нейрона.
Временная (последовательная) суммация — увеличение синаптиче-
ского притока по одному афферентному входу. Развивающиеся с ко-
ротким интервалом ВПСП суммируются, что вызывает пороговый
сдвиг МПП и генерацию ПД. Этот вид суммации наблюдается при уве-
личении силы раздражителя и возрастания частоты ПД в афферентных
волокнах.
Пространственная (одновременная) суммация — увеличение си-
наптического притока при одновременной активации нескольких си-
наптических входов. ВПСП возникают одновременно в большом коли-
честве близко расположенных синапсов. Этот вид суммации наблюда-
ется при увеличении рецептивного поля рефлекса, что приводит
к сокращению латентного времени рефлекса.
7. Явление облегчения — открыто Ч. Шеррингтоном. Является
следствием суммации возбуждения в нервных сетях. При одновремен-
ной стимуляции двух афферентных входов ответ больше, чем при раз-
дельной стимуляции. Это объясняется тем, что при раздельной стиму-
ляции входов часть из них являются подпороговыми, а при совместной
стимуляции ВПСП суммируются и МПП снижается до КУД.
8. Явление окклюзии (закупорки) — открыто Ч. Шеррингтоном. Яв-
ление обратное облегчению. При одновременной стимуляции несколь-
ких афферентных входов развивающийся ответ меньше ожидаемой
суммы их раздельной стимуляции. Эффект возникает вследствие кон-
вергенции афферентных входов на одних и тех же нейронах нервного
центра.
9. Трансформация ритма в нейронных сетях — несоответствие рит-
ма импульсации нейронов на входе в нервный центр и на выходе. Как
правило, на входе ритм нерегулярный (пачковый рисунок активности),
а на выходе из нервного центра ритм становится регулярным, характе-
ризуется примерно одинаковым межимпульсным интервалом. 10. Реверберация возбуждения (циркуляция) — ПД длительно цир-
кулирует по нейронным сетям, имеющим положительные обратные
связи. Циркуляция продолжается до тех пор, пока не разовьется утом-
ление или не включатся тормозные нейроны.
11. Последействие — следствие реверберации, сохранение активно-
сти в нервном центре после прекращения его стимуляции. Это явление
выражено в нейронах гиппокампа, полагают, что оно лежит в основе
формирования кратковременной памяти.
12. Габитуация — привыкание нервных сетей к действию повтор-
ных, монотонных, биологически незначимых раздражителей.
13. Синаптическая депрессия — утомление нервных сетей при их
длительной стимуляции. Может быть вызвана истощением запасов ме-
диатора, снижением чувствительности постсинаптической мембраны
к медиатору (десенситизация синапсов).
14. Пластичность — взаимозаменяемость нервных сетей. При нару-
шении одной нейронной сети ее функции может частично выполнять
другая сеть. Торможение — это нервный процесс, при котором возбуждение од-
них нейронов подавляет или предупреждает возбуждение других ней-
ронов. Торможение никогда не распространяется по нервным волокнам.
Это местный процесс. Для торможения в ЦНС имеются специальные
тормозные нейроны и тормозные синапсы. Некоторые медиаторы
в ЦНС оказывают только тормозное действие, например глицин,
ГАМК, но есть медиаторы, которые оказывают как тормозное, так
и возбуждающее действие, — ацетилхолин, глутамат.
Классификация торможения
1. Постсинаптическое, гиперполяризационное торможение связано
с формированием на мембране ТПСП. Постсинаптическая мембрана
при этом гиперполяризуется, возбудимость нейрона понижается, в ре-
зультате он или уменьшает частоту, или прекращает генерировать ПД.
В и д ы п о с т с и н а п т и ч е с к о г о т о р м о ж е н и я
а) центральное
Поступательное опережающее торможение по Сеченову: при раз-
дражении ретикулярной формации ствола мозга импульсы по ретику-
лоспинальному тракту поступают в спинной мозг и активируют тор-
мозные интернейроны, которые тормозят альфа-мотонейроны. Тормо-
жение альфа-мотонейронов проявляется в увеличении латентного
времени спинальных рефлексов.
б) Поступательное реципрокн торможение по Шеррингтону: при
активации рецепторов растяжения какой-либо мышцы возбуждаются ее
альфа-мотонейроны, при этом одновременно (реципрокно, сопряженно)
тормозятся альфа-мотонейроны мышцы антагониста. Например, при
возбуждении моторного ядра мышцы сгибателя реципрокно тормозится
моторное ядро мышцы разгибателя. Значение: автоматическое облегче-
ние работы сустава. в) Возвратное торможение по Реншоу: при увеличении частоты им-
пульсации альфа-мотонейрона колатераль его аксона возбуждает тор-
мозный интернейрон (клетка Реншоу). Тормозный нейрон выделяет
медиатор глицин, который, вызывая процесс гиперполяризации, снижа-
ет величину деполяризации сомы альфа-мотонейрона, что приводит
к понижению частоты ПД. Значение: обеспечивает сохранение средней
частоты импульсации альфа-мотонейрона, нейрон импульсирует ста-
бильно в течение длительного времени, обеспечивая поддержание по-
зного тонуса. Вещества стрихнин, столбнячный токсин блокируют ра-
боту глицинергических синапсов, что приводит к судорогам.
г) Латеральное (окружающее) торможение: при активации одного
нейрона одновременно активируются интернейроны, которые тормозят
рядом лежащие нейроны. Это торможение хорошо изучено в сенсорных
системах, предполагается, что оно обеспечивает контрастность воспри-
ятия сигнала.
2. Пресинаптическое деполяризационное торможение уменьшает
эффект действия на клетку возбуждающих синапсов (торможение на
входе). Морфологической основой его являются аксо-аксональные си-
напсы. Медиатор — ГАМК. При выделении ГАМК в пресинаптической
терминали возбуждающего синапса развивается длительная деполяри-
зация (до 100 мс), что приводит к уменьшению выделения возбуждаю-
щего медиатора и падению амплитуды ВПСП. Ионный механизм пре-
синаптического торможения до конца не выяснен. Предполагают, что
происходит повышение проницаемости для ионов хлора, но двигается
он согласно электрохимическому потенциалу из клетки.
-
Вегетативная нервная система (ВНС) — это часть нервной системы,
иннервирующая гладкие мышцы, железы, сердце.
ВНС делят на отделы: ПСНС — парасимпатическая нервная систе-
ма; СНС — симпатическая нервная система; МНС — метасимпатиче-
ская нервная система.
Работа СНС и ПСНС контролируется высшим вегетативным цент-
ром — гипоталамусом.
Вегетативная иннервация имеет 2-нейронный принцип строения.
Тело первого эфферентного нейрона — преганглионарного (пре-ГН) —
лежит в ЦНС. Его аксон (преганглионарное волокно) выходит из ЦНС
и идет к вегетативному ганглию. В ганглии возбуждение передается на
2-й эфферентный вегетативный нейрон — постганглионарный (пост-
ГН). Его аксон (постганглионарное волокно) покидает ганглий и иннер-
вирует эффекторный орган. Ганглии вегетативной нервной системы
являются периферическими нервными центрами (НЦ). Обладают всеми
свойствами НЦ — одностороннее проведение, суммация, дивергенция,
конвергенция, трансформация ритма и т. д.
особенности вегет ганглиев
1. Медиатором ганглиев является Ах, который связывается с ионо-
тропными никотин-чувствительными синаптическими холинорецепто-
рами (Н-хр).
2. В синапсах длительная синаптическая задержка.
3. Длительный ВПСП.
4. ПД имеет длительную следовую гиперполяризация; которая при-
водит к трансформации ритма возбуждения в виде снижения частоты
ПД в постганглионарных волокнах (не более 10—15 имп/с).
Симпатический отдел
Центры СНС локализованы в торако-люмбальном отделе спинного
мозга. Преганглионарные нейроны лежат в боковых рогах от последне- го шейного до 4-го поясничного сегмента спинного мозга (С8, Th1—
Th12, L1—L4).
Имеется 2 типа ганглиев в СНС. Паравертебральные ганглии явля-
ются парными и образуют симпатическую нервную цепочку по обе
стороны от спинного мозга (симпатический ствол). Превертебральные
ганглии — непарные, их три (солнечное сплетение, верхний и нижний
брыжеечные узлы).
Медиаторы реактивные системы
Преганглионарные нейроны — холинергические (Ах). Постганглио-
нарные нейроны — адренергические (НАд), передача возбуждения на
орган осуществляется с помощью альфа- и бета-адренорецепторов, ко-
торые являются метаботропными рецепторами.
При возбуждении альфа-адренорецепторов активируется мембран-
ный фермент фосфолипаза С и образуется два вторичных посредника:
инозитолтрифосфат (ИТФ) и диацилглицерол (ДАГ).
При возбуждении бета-адренорецепторов активируется мембранный
фермент аденилатциклаза, что приводит к образованию вторичного по-
средника ц-АМФ. Следствием активации адренорецепторов может быть
изменение как натриевой, так и калиевой проводимости мембраны эф-
фекторных клеток.
Парасимпат отдел
Центры ПСНС лежат в головном мозге (краниальный отдел). Это
средний мозг — ядро Якубовича (III пара черепномозговых нервов),
мост и продолговатый мозг — лицевой нерв (VII пара), языкоглоточ-
ный (IX пара), блуждающий нерв (X пара). В спинном мозге парасим-
патические центры локализованы в крестцовых сегментах (S2—S4).
Ганглии ПСНС располагаются около органа или внутри него, т. е.
интрамурально.
Медиаторы и реактивные системы
Преганглионарные нейроны — холинергические (Ах), синаптиче-
ские рецепторы — Н-холинорецепторы. Постганглионарные нейро-
ны — холинергические, рецепторы — М-холинорецепторы (мускари-
новые). М-холинорецепторы бывают двух видов: ионотропные и мета-
ботропные. В атипических кардиомиоцитах, некоторых кровеносных
сосудах взаимодействие Ах с М-холинорецепторами приводит к акти
метасимпатич отдел
Представляет собой внутриорганный отдел вегетативной системы.
Располагается в стенках внутренних органов. Контролирует относи-
тельно независимо от ЦНС внутренние органы на основе перифериче-
ских рефлекторных дуг. Регулирует автоматию внутренних органов
(сердце, кишечник), местный кровоток, секреторную и моторную
функции. В рефлекторной дуге имеются чувствительные, вставочные
и эффекторные нейроны. Медиаторы эффекторных нейронов: НАд, се-
ротонин, АТФ и др. Функциональное значение метасимпатической сис-
темы заключается в возможности саморегуляции органов независимо от
ЦНС, а также освобождение ЦНС от избытка информации.
1. Внутриорганный — метасимпатическая нервная система.
2. Спинальный — центры СНС и ПСНС.
3. Стволовой — в ретикулярной формации ствола мозга находится
сосудодвигательный центр, который контролирует работу СНС. Ядра
блуждающего нерва, лицевого, языкоглоточного, глазодвигательного
включают парасимпатические центры.
4. Гипоталамус — высший центр регуляции вегетативных функций.
Главный центр гомеостаза и формирования целенаправленных пове-
денческих реакций — биологических мотиваций.
5. Лимбическая система — участвует в адаптации организма к ме-
няющимся условиям окружающей среды, формировании мотивацион-
ного поведения и эмоций.
-
6. Лобная кора — обеспечивает контроль мотивационного поведения.