

специальная селекция
.pdfселекції врожайних сортів з високим умістом олії, стійкістю до вовчка, іржі та інших небезпечних хвороб і паразитів.
Використовуючи їх величезний досвід, селекціонери Інституту рослинництва ім. В.Я. Юр'єва (Вольф В.Г., Гуменюк А.Д.), Селек- ційно-генетичного інституту (Погорлецький Б.К.) досягли значних успіхів у створенні сортів-популяцій.
У 1974 році по Харківській області був розповсюджений середньостиглий сорт Харківський 100 з вегетаційним періодом 105110 діб, урожаєм насіння 2,17-2,55 т/га, збором олії 1040-1170 кг/га, вмістом олії 53-54 %.
Значний прогрес харківських селекціонерів було закладено в сорті соняшнику Харківський 50, який вирощувався з 1989 р. у Дніпропетровській, Донецькій і Харківській областях. Особливістю сорту була висока олійність насіння (до 59,5%) і скоростиглість (95-100 діб). У сорті Харківський 101, який було занесено до Реєстру сортів рослин України у 1980 р. по Луганській і Харківській областях, було також поєднано високу врожайність, скоростиглість і олійність, а також крупність насіння.
Селекція сортів-популяцій здійснювалася за схемою академіка B.C. Пустовойта, яка представлена на рис. 10.
Рис. 10. Схема селекції соняшнику академіка В. С. Пустовойта
230

Селекція сортів-популяцій, стійких до вовчка і несправжньої бо-
рошнистої роси. Селекційна робота щодо створення високоврожайних, стійких до основних патогенів сортів-популяцій соняшнику, повинна проводитися постійно, тому що внаслідок еволюції виникають нові вірулентні раси збудників хвороб.
Починаючи з 1982 року, в Інституті рослинництва ім. В. Я. Юр'єва УААН було розпочато реалізацію селекційної програми зі створення сортів соняшнику з груповою стійкістю до різних рас вовчка і несправжньої борошнистої роси. З метою швидкого виконання програми було залучено вихідний матеріал, отриманий за участю міжвидової гібридизації між дикими видами і культурним Helianthus annuus, створення якого почали з 1966 року В.Г. Вольф і М.С. Ситник.
Донорами стійкості використали гексаплоїдні і тетраплоїдні дикі види. Особливо цінними виявилися гексаплоїдні види, які відрізня-
лись груповим імунітетом до Orobanche cumana Wallr., Plasmopara helianti Novot., Puccinia helianti Szhw. Дикі види, за оригінальною схемою Г. В. Пустовойт, схрещували з кращими культурними сортами соняшнику ВНДІОК 6540 пол., Передовик, Старт, Харківський 100. В результаті насичувальних і парних схрещувань з багаторазовим індивідуальним добором, перевіркою нащадків на штучному фоні, інфікованому вовчком, було виділено велике різноманіття нових форм з різним ступенем стійкості (рис. 11).
Маса
Маса
Рис. 11. Схема селекції вихідного матеріалу – міжвидові гібриди
231
М.С. Ситник, А.Д. Гуменюк виділили декілька середньостиглих (МВГ-75, МВГ-136, МВГ-441, МВГ-755, МВГ-772); ранньостиглих (МВГ-551, МВГ-559, МВГ-762, МВГ-10) селекційних номерів з груповою стійкістю до основних хвороб і вовчка, які забезпечили створення сортів та ліній соняшнику.
11.10. Гетерозис і його практичне використання у соняшнику
Аналіз існуючих гіпотез і методологічної проблеми гетеро-
зисної селекції. Проблеми теорії селекції на сьогодні мають актуальність, а їх вирішення залежить від розробки методологічних питань. Це викликано, у першу чергу, вимогами до біологічних засобів виробництва, тобто сортів і гібридів сільськогосподарських культур.
Гетерозис – одне з імовірних явищ, притаманне багатоклітинним організмам, інтерес до якого проявляється сотні років. Прагнення до практичного використання його можна порівняти за часом з історією виникнення людської діяльності. Але, незважаючи на величезну зацікавленість з боку практичної селекції в необхідності теоретичного пояснення мікроеволюційних процесів у природних і експериментальних популяціях, все ж таки відсутня загальна теорія гетерозису.
Термін "гетерозис", введений Г. Шеллом, означає відхилення від адитивного ефекту батьків. Існуючі моделі (теорії) гетерозису пояснюють його природу з погляду знань, накопичених класичною генетикою на різних етапах її розвитку. Осмислювання природи гетерозису з позицій сучасного рівня знань біології щодо багатоклітинності організмів приводить до висновку про об'єктивну складність цього явища, неможливість пояснення його в термінах класичної дії і взаємодії генів.
Перше наукове пояснення гетерозису дав Ч. Дарвін, який розглядав статевий процес у рослин з погляду зміни організації принципів системних процесів на рівні макросистеми. Гетерозис він відносив до комплементарного прояву епігенетичних систем батьків у гібридах першого покоління.
В основу всіх моделей закладені дві концептуальні передумови. Перша полягає в постулюванні можливості пояснення природи гетерозису на основі закономірностей класичної дії і взаємодії генів. Друга – у тому, що акцентується увага на фактах щодо механізмів функціональної реалізації різноякісності і фізіологічній диференційованості батьківських форм.
232
Теорія генного балансу, теорія генетичного гомеостазу, теорія з погляду фізіологічних, біофізичних, біохімічних і молекулярногенетичних процесів повно представлені в роботах відомих вчених: К.Мазера, M.Lernera, Ц.М. Шерешевської, В.Г. Шахбазова, П.П. Літуна, М.П. Дубініна. В останній період широку популярність набула гіпотеза збалансованого метаболізму. Ця гіпотеза орієнтує на вивчення ферментних систем у батьківських форм і прогнозування на цій основі гетерозисних комбінацій. Одним із варіантів була гіпотеза доукомплектування ферментів Ф.Ф. Мацкова. В основу мо- лекулярно-генетичних гіпотез гетерозису покладені експериментальні факти міжалельної комплементації, що виявляється на рівні ізоферментів і біосинтезу інших речовин. В останнє десятиліття з'явилася можливість вивчення мінливості вихідного матеріалу і оцінювання генетичної близькості за допомогою генетичних маркерів
(RFLP, AFLP, SSR).
Методи вивчення генетичної цінності батьківських форм і прогнозування гетерозисних комбінацій. Оцінка генетичної і селек-
ційної цінності батьківських форм соняшнику має принципове значення для практичної селекції. При цьому результат залежить від вихідної теоретичної моделі гетерозису, на підвалинах якої розроблені конкретні методи оцінювання. Для практичної селекції важлива оптимальність оцінювання селекційної значимості вихідної форми і ефективність вибору гетерозисної комбінації.
Найбільш детальну розробку отримали методи оцінювання, які базуються на постулатах моделі полігенного контролю кількісних ознак. В їх основу покладено уявлення щодо можливості експериментального оцінювання двох компонентів генетичної цінності і загальної (ЗКЗ), і специфічної (СКЗ) комбінаційної здатності.
Розподіл поняття комбінаційної здатності на загальну і специфічну запропонували Г.Спрег та Л.Тетум. Вони розробили математичну модель оцінювання комбінаційної здатності. Удосконалення прийомів математичної моделі провів Б.Гріфінг.
Завдання оцінювання ліній на ЗКЗ складається з виявлення таких ліній, у яких під час схрещування спостерігається виділення переважних гібридних комбінацій над батьківськими формами і кращими гібридами. Оцінка на СКЗ проводиться з метою виділення конкретних пар ліній соняшнику, які забезпечують прояв гетерозису. Загальну комбінаційну здатність можна оцінити в різних системах схрещування: вільного запилення, полікросу, топкросу, діалельного схрещування. Специфічна здатність оцінюється лише в топкросних і діалельних схрещуваннях. Причому в топкрос для цих цілей застосовуються як тестери інбредні лінії соняшнику або прості гібриди з широкою генетичною основою.
233
За даними Л.В. Бондаренка, використання методу вільного запилення соняшнику давало кращі результати, ніж полікрос, і дозволило бракувати більше 40% досліджуваних ліній, які мали низьку ЗКЗ.
Вперше у 1932 році діалельні схрещування ліній соняшнику були проведені на Саратовській дослідній станції, у 1935 році – на Армавірському опорному пункті. Вивчення цього методу показало придатність його для селекційного оцінювання ліній соняшнику і виявлення високогетерозисних комбінацій.
Гетерозисна селекція соняшнику на якість продукції шляхом поліпшення складу жирних кислот. Молекулярна біологія, завдяки удосконаленню біохімічних лабораторій, зробила можливим вивчення процесів метаболізму – однієї з фундаментальних властивостей живих істот як відкритих систем.
У селекції соняшнику цікавими є знання щодо організації біогенезу сполук, які зумовлюють якість продукції. При цьому увага акцентується на інформації про місце і час біосинтезу вихідних продуктів у рослині, довжині окремих етапів, транспортуванні у місця біосинтезу запасних речовин у насіння, генетичній організації вторинного синтезу, а в цілому – на узгодженості всіх етапів і можливостях їх контролю.
Водночас біосинтез усього комплексу біохімічних компонентів, які беруть участь у синтезі жирних кислот на організмовому рівні, вивчено недостатньо. Можна з упевненістю засвідчити наступне:
-біосинтез пальмітинової кислоти (16:0) являє собою багатоступінчастий процес на базі вихідних сполук ацетил – КоА, який каталізується багатоферментним комплексом, детермінованим блоком структурних генів і здійснюється в пластидах клітини у рослин соняшнику;
-біосинтез стеаринової кислоти (18:0), як процесу перетворення 16:0 у 18:0, завдяки дії багатоферментного комплексу лігаз, що здійснює подовження вуглеводного ланцюга;
-біосинтез олеїнової кислоти, як процесу перетворення насиченої кислоти 18:0 у ненасичену багатоферментним комплексом десатураз I 9;
-біосинтезу лінолевої кислоти, як перетворення мононасиченої кислоти 18:1 у поліненасичену 18:2 багатоферментним комплексом десатураз I 12;
-біосинтезу ліноленової кислоти як перетворення полінасиченої 18:2 у 18:3 багатоферментним комплексом десатураз I 15.
Сьогодні вже відомо, що завдяки використанню мутагенів отримані форми соняшнику з умістом олеїнової кислоти 88-93 %, пальмітинової – 20-32, стеаринової – 8-12, лінолевої – 60-75 %. Вперше в Україні зареєстровані гібриди соняшнику з високим умістом олеїнової кислоти: Слав’янин, Дарій, Еней, Квін, Богун, Одол, і перший у світі пальмітиновий гібрид Капрал. Проводиться селекційна робота і випробування ліній та гібридів стеаринового, а також проміжних типів. Створення ліній і гібридів проводиться за схемою, яка представлена на рис. 12.
234
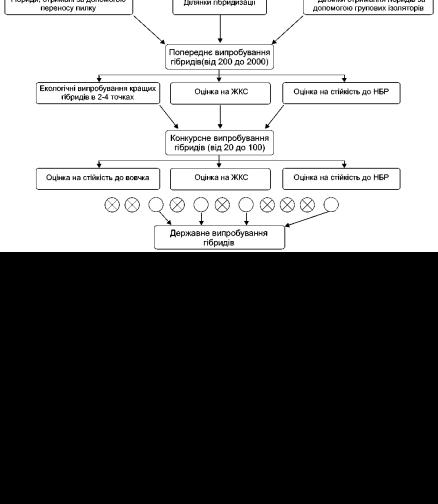
Рис. 12. Схема створення ліній та гібридів зі зміненим складом жирних кислот (Інститут рослинництва ім. В.Я. Юр'єва НААНУ)
11.11. Інбридинг в селекції соняшнику
Методи створення самозапилених (інбредних) ліній соняш-
нику. У соняшнику інбредні лінії отримують внаслідок примусового самозапилення окремих рослин сортів-популяцій, гібридів першого покоління, мутантів, синтетичних популяцій, міжвидових гібридів тощо.
235
У період на початку цвітіння селекціонери надівають на кращі корзинки пергаментні або тканинні ізолятори.
Після дозрівання насіння з кожної рослини збирають окремо. Як правило, з одного кошика можна отримати від 10 до 500 насінин залежно від вихідного матеріалу. Після досушування в сушарках за температур від 32 до 54°С насіння пакують.
Насіння з рослини першого самозапилення (І0) висівають в окремі рядки або на окремих ділянках. Самозапилення повторюють вибірково, залежно від завдань, які стоять перед селекціонером.
Рослини І1 відрізняються одна від одної тому, що відбувається розщеплення морфологічних і кількісних ознак батьків, які мають більшу гетерозиготність.
Уподальшому насіння соняшнику висівають індивідуально за
потомствами. В І2 помітна диференціація на окремі групи схожих рослин. В окремих потомствах виявляються слаборозвинуті рослини, які більш сприйнятливі до основних хвороб. Помітна різниця за висотою. Виявляються рослини з різним забарвленням язичкових квіток з їх різною щільністю, розміром тощо. Окремі рослини не здатні
продукувати пилок, особливо з F1, які створені з участю цитоплазма- тично-стерильних форм.
В І2 виявляються рослини соняшнику з різним типом галуження стебла, типом кошика, забарвленням листя. Всі рослини з ознаками депресії вибраковуються. З кращих нащадків добирають кращі рослини, які самозапилюють. Отримане насіння висівають індивідуально на окремих рядках (ділянках).
В І3 ще помітна диференціація, але є можливість відібрати однотипні рослини. В цьому поколінні інбридингу селекціонер має можливість відібрати велику колекцію ліній, які різняться окремими
ознаками. Окремі лінії в І3 мають стан стабілізації ознак, особливо з тих гібридних комбінацій, які мають однотипних батьків. Велика кількість інбридинг-поколінь бракується через сильну депресію. Чим більш гетерогенний вихідний матеріал, тим більше можна
отримати різноманітних ліній. Потомства І3 починають всебічно вивчати на комбінаційну здатність і стійкість до небезпечних хвороб та паразитів.
Уподальших поколіннях проводять постійний добір кращих рослин шляхом самозапилювання. Після 6-9 поколінь самозапилення і доборів лінії стають гомозиготними, вони відрізняються від вихідного матеріалу практично за всіма ознаками.
Оцінювання інбредних ліній на комбінаційну здатність. Ново-
створені лінії соняшнику в першу чергу необхідно перевірити на комбінаційну здатність. Визначити кращі лінії щодо загальної і специфічної комбінаційної здатності з урахуванням інших якісних гос-
236
подарсько-корисних ознак. Теоретично кожна лінія соняшника в схрещуваннях з іншими повинна показати ефект гетерозису. Перевага використання інбредних ліній у гетерозисній селекції в тому, що селекціонер має всі позитивні і негативні показники за ознаками в базі даних. Лінії можна отримувати безперервно в будь-якій кількості, з бажаними для гібриду F1 корисними ознаками. Але це не означає, що в практичній селекції всі гібридні комбінації є корисними у промисловому виробництві. Після ретельної перевірки по багатьох напрямах (прояв гетерозису, стійкість до основних хвороб, уміст олії, якість олії, толерантність до загущення, стійкість до гербіцидів, висота рослини, продуктивність пилку у відновників фертильності та її життєздатність, урожайність материнських форм, стійкість до вилягання та ін.) приймається рішення про подальшу стратегію використання створених інбредних ліній.
Комбінаційну здатність (КЗ) неможливо оцінювати візуально або непрямими методами у зв'язку з тим, що ефекти її коливаються залежно від пар схрещування, місця вирощування, року та ін. Існує єдиний шлях перевірки ліній стосовно КЗ – це схрещування з наступним оцінюванням F1, як правило, впродовж 3-х років, бажано в декількох географічних зонах за єдиною технологією і методикою випробування.
КЗ ліній соняшника вивчають методами топкросу, полікросу, вільного запилення та в системі діалельних схрещувань як по повній, так і неповній схемах. У практичній селекції використовують метод топкросів, у генетичних дослідженнях – повні діалельні схрещування, які вимагають величезного обсягу робіт. Селекціонера насамперед цікавлять можливості використання материнських ліній та їх стерильних аналогів у промисловому виробництві. Материнським формам, перш за все, пред’являють вимоги щодо високого рівня ефектів специфічної комбінаційної здатності, тобто прояву ефекта гетерозису в конкретній комбінації. Крім того, існує великий перелік інших вимог, які будуть розглянуті в наступних розділах.
Основною вимогою з боку КЗ до відновників фертильності є високий рівень ефектів загальної комбінаційної здатності через те, що окремі лінії-відновники фертильності використовують у промисловому насінництві з багатьма материнськими формами. Наприклад, гібриди Еней, Квін, Дарій, Богун, Ант, Зорепад мають різні материнські форми і одну лінію-відновника фертильності пилку Х-526В; Оскіл, Романс, Ясон, Боєць - Х720В; гібриди Харківський 49, Світоч, Погляд, Харківський 58 – лінію Х711В.
Для оцінювання ефектів КЗ використовують математичні програми обчислювання, які представлені в спеціальній літературі. Система селекції гібридів соняшнику в Інституті рослинництва представлена на рис. 13.
237
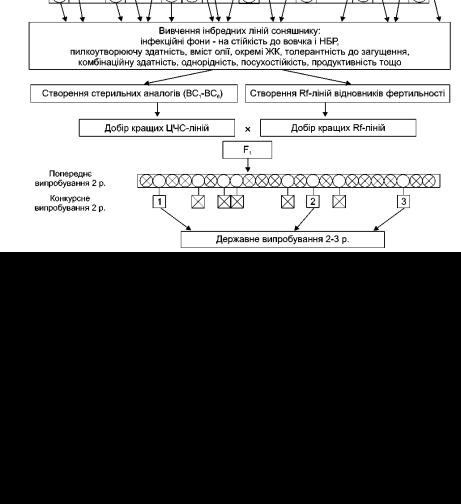
Рис. 13. Науково обґрунтована система селекції гібридів соняшнику
(Інститут рослинництва ім. В.Я. Юр'єва НААНУ)
Таким чином, з величезного матеріалу десятки тисяч гібридів першого покоління селекціонер відбирає близько 10 гібридних комбінацій для подальшого вивчення і впровадження у сільськогосподарське виробництво.
Оцінювання інбредних ліній на стійкість до вовчка. Небезпеч-
ний супутник соняшнику –вовчок (Orobanche cumana W.) – рослинапаразит, яка розмножується дрібним насінням, що утворюється на квітоніжці після перехресного запилення. Вовчок прижився на великій території Середземномор'я та Причорномор’я, але повністю від-
238

сутній на Американському континенті. За короткий час вовчок перетворився з раси А на расу Н, яка лютує в Іспанії, Туреччині і Болгарії. В Україні вовчок добре пристосувався до кліматичних умов Одеської, Херсонської Запорізької, Донецької і Харківської областей (південні регіони). Таким чином, одним із завдань селекції соняшнику є постійний моніторинг розповсюдження вовчка та створення нових ліній, стійких до існуючих і нових рас паразита.
Оцінювання нового селекційного матеріалу здійснюється як у кліматичній камері (взимку), так і в польових умовах за штучного зараження (рис. 14). Внесення насіння вовчка здійснюється як у вазони, так і в кожне гніздо разом з посівом соняшнику по 2 г на одну рослину.
Рис. 14. Лабораторний (а, б) та польовий (в) методи випробування стійкості сортозразків соняшнику до вовчка
Оцінювання інбредних ліній на стійкість до несправжньої боро-
шнистої роси. Збудник несправжньої борошнистої роси (Plasmopaza helianthi Novot.) є облігатним паразитом і розвивається тільки на живих рослинах. На сьогодні селекціонерам відомо 12 фізіологічних рас. Несправжня борошниста роса зустрічається в усіх зонах вирощування соняшнику, але найбільш поширена в Лісостепу і північній частині Степу. Недобір урожаю від шкоди цього патогена може складати до 50 %.
Хвороба діагностується впродовж всієї вегетації рослин – від сходів до повної стиглості і проявляється у двох формах залежно від періоду зараження і фази розвитку соняшнику:
-за первинного ураження – у вигляді загального пригнічення (дифузне ураження);
-за вторинного ураження з’являються плямистості листя (локальне або місцеве ураження).
Первинне враження рослин відбувається через кореневі волоски внаслідок проникнення в них гриба під час проростання зооспор у кореневу систему проростків. Далі гриб розвивається всередині рослини міжклітинно, піднімається за точкою росту вгору до наземних органів (дифузне враження). Критичним періодом для цього є фаза від проростання насіння до першої пари справжніх листків.
239