

специальная селекция
.pdfний ген інших видів, що контролює не притаманні вихідній формі ознаки (наприклад, стійкість до гербіцидів певного типу), форми з округлим коренеплодом, з покращеним метаболізмом ( оптимізованим фотосинтезом і розподілом цукрів у коренеплоді), самофертильні (за примусового запилення формують насіння).
Кожна з цих форм може бути представником будь-якого екотипу, проте всі вони характеризуються специфічними екологічними, морфологічними, цитологічними і генетичними особливостями, що визначають різновидності цукрових буряків.
Вихідний матеріал для селекції цукрових буряків. Вихідним ма-
теріалом у селекції є культурні й дикі види і різновидності, сортипопуляції, продукти масових та індивідуально-родинних доборів, гібридні міжлінійні, сортолінійні та міжсортові зразки, лінії різних інбредних поколінь, синтетики різних циклів періодичних доборів.
Потребам сучасної селекції буряків слугують також зразки Національного центру генетичних ресурсів рослин України, який діє на базі Інституту рослинництва ім. В.Я. Юр’єва (м. Харків), що налічує більше 124 тис. зразків, у тому числі 5,6 тис. технічних і 2,9 тис. кормових культур. Крім того, колекція Інституту цукрових буряків та його мережі нараховує біля 300 зразків, які репрезентують 12 видів і щорічно поповнюється ознаковими донорами, новими культурними формами й дикими співродичами.
14.2. Лінійна селекція цукрових буряків
Інбридинг і його вплив на цукровий буряк. Основним методом одер-
жання ліній у цукрових буряків є інбридинг, який у сучасних генетикоселекційних дослідженнях оцінений по-новому, його значущість очевидна як у теоретичних розробках, так і на практиці. Якщо основою популяційної селекції були різні форми масового та групового доборів, то у гетерозисній селекції необхідним є використання ліній, гомозиготних за селектованими ознаками, і їх наступна гібридизація.
Шкідливий вплив самозапилення на цукрові буряки вивчали В. Бартос, К. Андрлік, Г. Феліх, Г. Корн, Б. Каянус, Г. Шоу, Е. Бауер. Всі вони відмічали, що за примусового самозапилення зав’язування насіння погіршується, з’являються дефективні рослини, знижуються показники схожості насіння, врожайності і цукристості коренеплодів (депресія). Створені таким чином лінії селекційного значення не мали, проте метод інбридингу дозволив провести диференціацію складної перехреснозапильної популяції і виділити в гомозиготному стані багато відмінних одна від одної ліній, а також виділити цінні ознаки, що мають рецесивну природу. Тому інбридинг М.І. Вавілов справедливо вважав формотворчим фактором. Це дозволяє розкласти популяції вихідних матеріалів буряків на ботанічні форми (однонасінні, самофертильні та стерильні за пилком і т.п.).
280
Створенню самозапилених ліній перешкоджає система генетичної несумісності, яка притаманна перехреснозапильним культурам, у тому числі й цукровим бурякам. Для цієї культури характерні такі типи запилення: перехресне запилення між квітками різних рослин (ксеногамія), запилення між квітками однієї й тієї ж рослини (гейтеногамія) і самозапилення у межах однієї квітки (аутогамія). Проте окремі рослини у межах популяції здатні до самозапилення, що вперше довів Ч. Дарвін. Проте пізніше Hjalmar-Nilson, Hallgvist вказали на обмеженість самозапилення у цукрових буряків.
Генетична система самонесумісності пояснюється гіпотезою опозиційних факторів. У цукрового буряку вона належить до гаметофітного типу, особливістю якої є незалежна дія множинної серії S-алелів у пилкових зернах і приймочці маточки. Генетичну інтерпретацію самонесумісності дає гіпотеза Оуена (1942), згідно з якою ця ознака контролюється двома комплементарними генами, що виявляють незалежну дію один до одного. За наявності однакових алелів у пилковій трубці і тканині приймочки рослина не здатна до самозапилення, проте зав’язує насіння від запилення чужорідним пилком, який несуть неідентичні алелі. Так, якщо рослина має генотип s1s2s3s3, то пилкові трубки генотипу s1s3 і s2s3 викликають реакцію несумісності, натомість, попадання пилку, що несе алелі s1s4, s2s4, s1s5, s2s6 і т.д., цю реакцію пригнічує, внаслідок чого на материнській рослині формується насіння. Відмінності між генотипами пилкового зерна і приймочки хоча б за одним алелем приводять до самозапилення. Дигенний контроль ознаки самонесумісності підтверджено й іншими дослідниками.
У нашій країні дослідження з інцухту цукрових буряків було вперше проведено Т.Ф. Гриньком на Іванівській дослідно-селекційній станції. Популяції цукрових буряків представлені в основному самостерильними рослинами, проте відмічені рослини, що зав’язують невелику кількість насіння. Специфічна реакція на самозапилення у популяцій цукрових буряків має генетичну зумовленість. Різна кількість рослин, що зав’язали насіння, пояснюється генетичним різноманіттям вихідних матеріалів, на яких закладалися лінії, а також модифікаційним впливом довкілля.
Проблема інцухту не зводиться до виділення самофертильних біотипів, значення цього методу слід розглядати ширше. Воно полягає у перебудові генотипної структури перехреснозапильних популяцій. Генетичні процеси, які відбуваються під час самозапилення, призводять до того, що гени переходять у гомозиготний стан, і це викликає депресію майже всіх кількісних ознак. Негативний вплив інбридингу спричиняється тим, що шкідливі рецесивні мутації, пригнічені домінантними алелями у гетерозиготному стані, переходять у гомозиготний стан і проявляються на фенотипічному рівні зниженими показниками життєздатності, врожайності, стійкості до хвороб. В Інституті цукрових буряків одержано експериментальні дані щодо впливу інбридингу на госпо-
281
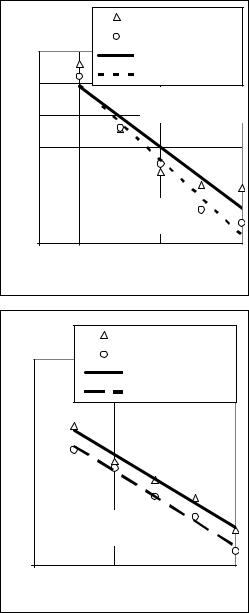
дарсько цінні ознаки цукрових буряків (популяцій урожайного і цукри- |
||||||||||||||||||
стого напрямів доборів та інбредних ліній чотирьох інбредних поколінь |
||||||||||||||||||
інбридингу). За однакових |
|
|
|
|
|
|
|
|
|
|||||||||
вихідних |
фенотипів |
ці |
|
|
|
|
В 1002 |
|
|
|
|
|||||||
ознаки знижувалися в обох |
|
|
|
|
ЛР 14759 |
|
|
|
|
|||||||||
зразках з різною інтенсив- |
|
|
|
|
|
|
|
|
||||||||||
|
5 |
|
|
Лінійний |
|
|
|
|
||||||||||
ністю (рис. 21 і рис. 22). |
|
|
|
|
|
Линейный (В 1002) |
||||||||||||
|
|
|
|
|
Лінійний |
|
|
|
|
|||||||||
У результаті послідов- |
|
|
|
|
|
|
|
|
||||||||||
|
4,5 |
|
|
Линейный (ЛР 14759) |
||||||||||||||
них |
поколінь |
інбридингу |
|
|
|
|
|
|
|
|
||||||||
|
|
|
|
y = -0,4758x + 4,9288 |
||||||||||||||
значення кількісних ознак |
/рослину |
|
|
|
r = - 0,92 |
R |
2 |
= 0,84 |
||||||||||
4 |
|
|
|
|||||||||||||||
перестає |
|
знижуватися, |
|
|
|
|||||||||||||
|
|
|
|
|
|
|
|
|
||||||||||
тобто |
настає |
|
інбредний |
3,5 |
|
|
|
|
|
|
|
|||||||
мінімум, іноді й після де- |
|
|
|
|
|
|
|
|||||||||||
2 |
|
|
|
|
|
|
|
|||||||||||
|
|
|
|
|
|
|
|
|||||||||||
. см |
|
|
|
|
|
|
|
|
||||||||||
сяти |
|
генерацій, |
що свід- |
|
|
|
|
|
|
|
|
|||||||
|
Тис |
|
|
|
|
|
|
|
|
|||||||||
чить про високу стабіль- |
3 |
|
|
|
|
|
|
|
||||||||||
|
|
|
|
|
|
|
|
|||||||||||
|
|
|
|
|
|
|
|
|
||||||||||
ність |
|
кількісних |
ознак у |
|
|
y = -0,5879x + 5,0677 |
|
|
|
|
||||||||
послідовних |
|
поколіннях |
|
2,5 |
|
|
|
|
||||||||||
|
|
|
|
|
|
|
|
|
||||||||||
|
|
|
r = - 0,98 |
R 2 = 0,97 |
|
|
|
|
||||||||||
самозапилених |
ліній. Де- |
|
|
|
|
|
|
|
|
|
||||||||
пресія за основними гос- |
|
2 |
|
|
|
|
|
|
|
|||||||||
|
0 |
1 |
2 |
3 |
|
4 |
5 |
|||||||||||
подарськими |
|
ознаками |
|
|
||||||||||||||
|
|
|
|
Покоління |
|
|
|
|
||||||||||
проявляється |
у |
цукрових |
|
|
|
|
|
|
|
|||||||||
|
|
A. Площа листкової поверхні |
||||||||||||||||
буряків з першого інбред- |
|
|
|
|
|
|
|
|
|
|||||||||
ного покоління, на що |
|
|
|
|
|
|
|
|
|
|||||||||
вказували |
майже |
всі |
до- |
|
|
|
В 1002 |
|
|
|
|
|||||||
слідники, які працювали з |
|
|
|
|
|
|
|
|||||||||||
|
|
|
|
|
|
|
|
|
||||||||||
самозапиленими |
лініями. |
|
70 |
|
ЛР 14759 |
|
|
|
|
|||||||||
Проте були випадки, коли |
|
|
|
Лінійний |
|
|
|
|
||||||||||
|
|
|
Линейный (В 1002) |
|||||||||||||||
не спостерігалася |
велика |
|
|
|
Лінійний |
|
|
|
|
|||||||||
депресія урожайності, цу- |
|
|
|
Линейный (ЛР 14759) |
||||||||||||||
|
|
|
|
|
|
|
|
|
||||||||||
кристості |
і |
габітусу рос- |
|
|
|
|
y = -3,57x + 63,35 |
|||||||||||
лин цукрових буряків, або |
|
60 |
|
|
|
2 |
|
|
|
|||||||||
ділянку |
|
|
|
r = - 0,99 R |
= 0,98 |
|||||||||||||
ж її зовсім не було. |
|
|
|
|
|
|||||||||||||
|
|
|
|
|
|
|
|
|
||||||||||
Відомо, |
що |
величина |
|
|
|
|
|
|
|
|
||||||||
інбредної |
депресії |
під |
са- |
|
|
|
|
|
|
|
|
|||||||
Кг/ |
|
|
|
|
|
|
|
|
||||||||||
мозапилення |
|
у |
диплоїдів |
|
|
|
|
|
|
|
|
|||||||
|
|
50 |
|
|
|
|
|
|
|
|||||||||
змінюється, як правило, від- |
|
|
y = -3,67x + 61,09 |
|
|
|
|
|||||||||||
повідно до коефіцієнта ін- |
|
|
|
|
|
|
||||||||||||
|
|
r = - 0,99 R 2 |
= 0,99 |
|
|
|
|
|||||||||||
бридингу (F), який показує |
|
|
|
|
|
|
||||||||||||
|
|
|
|
|
|
|
|
|
||||||||||
ймовірність знаходження у |
|
40 |
|
|
|
|
|
|
|
|||||||||
гомологічних |
хромосомах |
|
|
|
|
|
|
|
|
|||||||||
|
0 |
1 |
2 |
3 |
4 |
|
|
5 |
||||||||||
двох ідентичних алелів ге- |
|
|
Покоління |
|
|
|
|
|||||||||||
на. Цей коефіцієнт уперше |
|
|
Б. Врожайність коренеплодів |
|||||||||||||||
був |
введений |
|
S.Wrigt |
у |
|
Рис. 21. Вплив інбридингу на ріст, |
||||||||||||
1924р. За самозапилення |
|
|||||||||||||||||
розвиток і врожайність коренеплодів |
||||||||||||||||||
диплоїдів |
F=1/2 |
(1+F1), |
де |
|||||||||||||||
|
|
|
|
|
|
|
|
|
|
282 |
|
|
|
|
|
|
|
|
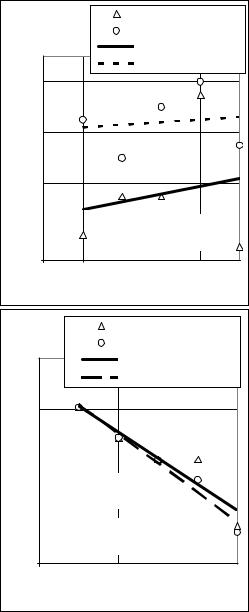
F1 – коефіцієнт інбридингу попереднього покоління. У диплоїдів гомо- |
|||||||||||||||
зиготизація наступає значно швидше, ніж у тетраплоїдів. Високий сту- |
|||||||||||||||
|
|
|
В 1002 |
|
|
пінь гетерозиготності |
але- |
||||||||
|
|
|
|
|
лів у тетраплоїдів пояснює |
||||||||||
|
|
|
ЛР 14759 |
|
причини їх могутності, крім |
||||||||||
|
|
|
Лінійний |
|
того, у них існує більш ши- |
||||||||||
|
16,4 |
|
Линейный (В 1002) |
|
|||||||||||
|
|
Лінійний |
|
рокий |
спектр |
алельних і |
|||||||||
|
|
|
|
||||||||||||
|
|
|
Линейный (ЛР 14759) |
||||||||||||
|
16,2 |
|
|
|
|
|
міжалельних взаємодій. |
||||||||
|
|
|
|
|
|
|
Інбридинг у гетерозис- |
||||||||
|
16 |
|
|
|
|
|
ній |
селекції |
|
необхідно |
|||||
|
|
|
|
|
|
|
|
||||||||
% |
15,8 |
|
|
|
|
|
застосовувати ще й тому, |
||||||||
Цукристість, |
|
|
|
|
|
що вже на перших етапах |
|||||||||
|
|
y = 0,02x + 15,82 |
|
||||||||||||
15,6 |
|
r = 0,14 R 2 |
= 0,02 |
|
селекційної роботи елімі- |
||||||||||
|
|
|
|
|
|
нуються дефективні і не- |
|||||||||
15,4 |
|
|
|
|
|
життєздатні |
рослини, які |
||||||||
|
|
|
|
|
|
||||||||||
|
|
|
|
|
|
несуть летальні і напівле- |
|||||||||
|
15,2 |
|
|
|
|
|
|||||||||
|
|
|
|
|
|
тальні гени. |
Переведення |
||||||||
|
|
|
y = 0,06x + 15,14 |
|
|||||||||||
|
|
|
|
мутації у |
гетерозиготний |
||||||||||
|
15 |
|
r = 0,2 R 2 = 0,04 |
|
|||||||||||
|
|
|
|
||||||||||||
|
|
|
|
|
|
|
стан |
дозволяє |
компенсу- |
||||||
|
14,8 |
|
|
|
|
|
вати їх шкідливий вплив і |
||||||||
|
0 |
1 |
2 |
3 |
4 |
5 |
|||||||||
|
забезпечує високий рівень |
||||||||||||||
|
|
|
Покоління |
|
|
||||||||||
|
|
В. Цукристість коренеплодів |
|
продуктивності та життє- |
|||||||||||
|
|
|
|
|
|
|
здатності гібридів за уча- |
||||||||
|
|
|
В 1002 |
|
|
стю таких форм. |
|
|
|
||||||
|
|
|
ЛР 14759 |
|
|
Зняття |
інбредної |
де- |
|||||||
|
10,0 |
|
Лінійний |
|
|
пресії |
можливе |
за |
умови |
||||||
|
|
|
Линейный (В 1002) |
|
гібридизації |
контрастних |
|||||||||
|
|
|
ЛінійнийЛинейный (ЛР 14759) |
|
|||||||||||
|
|
|
|
за ознаками ліній. Для цук- |
|||||||||||
|
|
|
|
|
|
|
|||||||||
|
9,0 |
|
|
|
|
|
рових буряків лінії-запи- |
||||||||
|
|
|
|
|
|
|
лювачі схрещують з чоло- |
||||||||
Кг/ділянку |
|
|
|
|
|
|
вічостерильними формами, |
||||||||
|
|
|
|
|
|
причому цінність цих ма- |
|||||||||
8,0 |
|
|
|
|
|
теринських |
ліній |
також |
|||||||
|
|
|
|
|
|
||||||||||
|
y = -0,5062x + 9,58 |
|
|
|
залежить від генотипу. |
|
|||||||||
|
|
r = - 0,94 R 2 = 0,88 |
|
|
|
Відомо, |
що |
процеси |
|||||||
|
7,0 |
|
|
|
|
|
формування ознак продук- |
||||||||
|
|
|
|
|
|
|
|||||||||
|
|
|
y = -0,57x + 9,6616 |
|
тивності цукрових буряків |
||||||||||
|
|
|
r = - 98 |
R 2 |
= 0,97 |
|
проходять |
в |
онтогенезі і |
||||||
|
6,0 |
|
|
|
|
|
залежать |
від |
особливос- |
||||||
|
0 |
1 |
2 |
3 |
4 |
5 |
тей розвитку рослин, пе- |
||||||||
|
|
|
|
|
|
|
|||||||||
|
|
|
Покоління |
|
|
ребігу фізіологічних |
про- |
||||||||
|
|
|
Г. Збір цукру |
|
цесів, взаємозв’язків між |
||||||||||
|
|
|
|
|
|
|
|||||||||
Рис. 22. Вплив інбридингу на основні |
врожайністю й цукристіс- |
||||||||||||||
тю, площею і станом лист- |
|||||||||||||||
господарсько цінні ознаки запилювачів |
|||||||||||||||
кового апарату. |
На |
|
про- |
||||||||||||
|
|
|
|
|
283 |
|
|
|
|
|
|
|
|
|
|
дуктивність також впливає й підсумкова листкова поверхня рослин. Для утворення маси коренеплоду істотне значення має величина асиміляційного апарату, а для цукристості, крім цього, і якість листкової тканини. Більш високу цукристість мають рослини з меншою кількістю відносно великих листків. Інші дослідники вважають, що цукристість коренеплодів більшою мірою, ніж урожайність, залежить від облистяності рослин. Особливу роль при цьому відіграють крупні листки другого десятка, які є найбільш продуктивними щодо формування врожаю і цукронакопичення. Проте має значення і зворотний фотосинтезу процес дихання, який пов’язаний із витратою накопичених органічних речовин.
Формування врожаю відбувається не лише на тлі складних взаємозв’язків і взаємозумовленості найважливіших фізіологічних процесів, але й залежить від спадкових особливостей генотипу, їх реакції на умови середовища (забезпеченість рослин вологою, ріст і розвиток за певного теплового режиму і т.п.). Іншими словами, це складний процес, на який впливає багато факторів.
У гетерозисній селекції під час створення ліній слід враховувати не лише ті показники, які формують продуктивність, а й репродуктивну здатність фертильних ліній-запилювачів, від якої залежить ефективність перезапилення, запліднення і в кінцевому результаті врожай та якість гібридного насіння. Запилення відіграє певну роль у формуванні різноманіття генотипів у популяції, робить ефективним добір, позитивно корелює зі ступенем зв’язування плодів і схожістю насіння, а величина пилкових зерен партнерів схрещування впливає на рівень гібридизації й вибірковість запліднення. Чим більша різниця між величиною пилкових зерен у батьків, тим вищим буде рівень гібридизації у форми з крупнішим пилком і нижчим – у більш дрібноклітинної форми.
За період цвітіння на кожну приймочку попадає не менше 300400 пилкових зерен. Цукровий буряк є анемофільною культурою, тому в процесі розмноження кількість і якість пилку набуває великого значення. Наявність великої кількості пилку створює конкуренцію гамет, оскільки добір може проходити як на рівні фенотипу, генотипу, зиготи, так і в гаплоїдній фазі.
Самозапилення рослин цукрових буряків, яке використовується у процесі створення лінійного селекційного матеріалу, неминуче пов’язане з його негативним впливом на процеси запліднення, ембріогенезу та формування життєздатного насіння, причому інцухтдепресія у разі зниження рівня гетерозиготності спостерігається незалежно від умов року і місця вирощування. Навіть однократне інцухтування знижує кількість пилкових зерен у пиляках майже вдвічі. За наступного інцухтування загальна тенденція зберігається, хоча й меншою мірою. За самозапилення відмічено порушення процесу мейозу як у цукрових буряків, так і в інших культур. Аномалії в процесі мейозу виявлені незалежно від сорту і ступеня інцухту.
284
Інбридинг призводить також до зменшення кількості нормальних і збільшення кількості карликових та дрібних пилкових зерен. Відмічена ще й тенденція зниження фертильності пилку з поглибленням інбридингу. Окремі рослини утворювали повністю стерильний пилок.
Запилення тісно пов’язане зі схожістю насіння. Досліджуючи кореляційні зв’язки між деякими ознаками насіннєвих рослин різних форм цукрових буряків, встановили середні позитивні кореляції між фертильністю пилку і схожістю насіння, а також зі ступенем зав’язування плодів.
Дослідники, які вивчали схожість насіння, вважають, що ця ознака має спадкову зумовленість, хоча значною мірою залежить і від неспадкових факторів. Ступінь гомозиготності теж впливає на ці показники. Встановлено, що саме гомозиготи та моногетерозиготи гинуть на різних етапах ембріогенезу або утворюють виповнені, але нежиттєздатні плоди. З підвищенням ступеня інбридингу диференціація і рівень схожості знижуються. Низька схожість насіння спричинена тим, що в результаті дії системи самонесумісності зародок дегенерує, зовні нормальні плоди є нежиттєздатними. Якщо рівень схожості насіння компонентів був невисоким (55-65%), то він суттєво підвищується під час відтворення гібридів у фабричній генерації. За даними іноземних авторів, схожість у потомстві від трьох послідовних самозапилень була нижчою, ніж у контролі, і її відновленню сприяло перезапилення інбредних ліній.
Таким чином, внаслідок інбредної депресії за багатьма господарсько цінними ознаками, що спостерігається у самозапилених поколіннях, гомозиготні лінії у практичній селекції самостійного значення не мають, тобто не можуть служити сортами. У генетичних дослідженнях такі лінії використовують для вивчення генетичної природи ознак, вони можуть бути їх донорами, а також сприяють виявленню цінних властивостей, що контролюються рецесивними генами (наприклад, однонасінність). Метод інцухтування має велике значення з точки зору наступної гібридизації ліній для одержання гетерозисних гібридів.
Методи підвищення результативності створення ліній. Врахо-
вуючи труднощі отримання насіння під час самозапилення, а також депресію основних ознак, яка викликає випадіння із селекційної роботи багатьох ліній, що досягли інбредного мінімуму, деякі дослідники перевагу віддають не суворому інбридингу, а так званому помірному інбридингу, за якого використовують сибси, напівсибси, інбредні популяції. Проте гетерозисний ефект гібридизації такого матеріалу значно нижчий, ніж від гібридизації самозапилених ліній. До інших способів підвищення результативності створення ліній слід віднести використання явища псевдосамофертильності. Відомо, що за рахунок понижених температур під час цвітіння, які можна досягти
285
як в умовах селекційно-тепличних комплексів, так і в природних (умови високогір’я), реакція самонесумісності пригнічується і тоді підвищується зав’язуваність насіння. Добір та закладення ліній можливі також і на самофертильних біотипах, проте частота гена самофертильності у природних популяціях є надто низькою. Виходом із цього може слугувати передача гена самофертильності самонесумісним формам від донорів цієї ознаки.
Сучасний стан генетико-селекційних досліджень вказує на те, що створення колекцій гомозиготних ліній цукрових буряків за певними ознаками – це перспективний шлях і важливий етап у гетерозисній селекції культури.
Способи ізоляції рослин. Для отримання ліній необхідна ізоляція рослин від чужорідного перезапилення. Для цього використовують індивідуальні й групові ізолятори різних конструкцій, що складаються з металевих конструкцій (каркасів) і бязевих чохлів. За конструкцією вони можуть бути різні – у вигляді ізоляційних “будиночків” та циліндричні, а також застосовують і безкаркасні пергаментні ізолятори.
Групові ізолятори використовують для одержання насіння за суворої ізоляції від невеликих груп рослин: запилювачів, закріплювачів стерильності О типу і їх чоловічостерильних (ЧС) аналогів, потомств самозапилених ліній та інших селекційних матеріалів, що мають цінні ознаки (підвищену цукристість, стійкість до хвороб, високу комбінаційну здатність і т.п.). Їх можна використовувати також для одержання насіння простих чоловічостерильних гібридів, отриманих для вивчення окремих ознак. За технічними можливостями під групові ізолятори виділяється спеціально обладнана стаціонарна ізоляторна ділянка, розташована на рівній місцевості. Якщо групові ізолятори більш складної конструкції мають, крім каркаса і чохла, ще й вентилятор, то ізоляторна ділянка повинна бути забезпечена водопровідною (для поливу) та електричною (для роботи вентиляторів) мережами. У групові ізолятори циліндричного типу висаджують коренеплоди концентричними колами. Зовнішнє коло поділяють на 14-16 частин, а внутрішнє – на 6-8. У разі розміщення під ізолятором двох номерів (наприклад, ЧС аналога і закріплювача стерильності) один номер висаджують по зовнішньому колу, а інший – по внутрішньому і в центрі ізолятора. На номері розміщують етикетку. Оптимальним строком установки металічних каркасів на ділянках насінників є період масової появи квітконосних пагонів. У період бутонізації, за два-чотири дні до початку цвітіння, проводять попереднє вибракування насінників за однонасінністю, стерильністю та іншими ознаками, а також обробку рослин проти хвороб і шкідників, потім на каркаси одягають чохли. Перед дозріванням насінників їх знімають, розділяють пагони різних номерів, підв’язують окремі рослини. У фазі повної стиглості насінники зрі-
286
зають за компонентами, підсушують, обмолочують, проводять первинне очищення насіння і враховують його масу.
Індивідуальні ізолятори слугують для ізоляції одиночних рослин, або ж для парних схрещувань. Якщо в індивідуальний ізолятор висаджують одну фертильну, а іншу – стерильну рослину, то насіння з фертильної рослини є першим інбредним поколінням, а зі стерильної
–гібридним, яке наступного року випробовується. Якщо в ізоляторі
–дві фертильні рослини, то насіння збирається з кожної рослини окремо. Ці гібриди є реципрокними, і такі схрещування використовують для рекомбігенезу ознак.
Використовують також і пергаментні, і целофанові безкаркасні ізолятори, проте головною умовою є вчасна їх постановка для перешкоджання неконтрольованого запилення.
14.3. Селекція на гетерозис на основі цитоплазматичної чоловічої стерильності
Теорії та теоретичні основи гетерозису. Першим дослідником,
який ще в середині 19-го століття зробив узагальнювальний висновок про виявлення гібридної сили за дії перехресного запилення, порівняно із самозапиленням у межах одного і того ж виду, був Ч. Дарвін, хоча перші наукові досліди зі схрещування різних видів тютюну та переваги щодо гібридних форм проведені ще раніше – у другій половині ХVIII ст. і належать І. Кельрейтеру. Із зародженням селекції як науки у цукрових буряків підвищення врожаю за схрещування у першому поколінні помічав ще Л. Вільморен, про що описано в “Історії фірми Вільморенів” П.Ф. Шліппе у 1929 р. Результати робіт зі схрещування різних рас цукрових буряків того часу опубліковано також і вітчизняними вченими С.В. Гудвілом та І.І. Войткевичем.
Генетичні дослідження цього явища, названого у 1914 році гетерозисом, були започатковані Шеллом, Істом, Джонсоном та іншими генетиками на початку ΧΧ ст. Значну частину наукових фактів, пов’язаних з гетерозисним ефектом, пояснювали як вплив добросприятливих домінантних алелів Давенпорт, Джонс, Колінз, пізніше Брус, Кібл, Пеллю, Річі, Спрег. Проте деякі факти, особливо “моноgene Heterosis”, які не можна було пояснити з позиції теорії домінування, підтверджували наявність наддомінування як наслідок дії гетерозиготних пар алелів. К. Мазер (1955), враховуючи цитоплазматичні особливості батьківських форм, причиною гетерозису вважав сумарну взаємодоповнюючу дію всіх елементів клітини, що зумовлюють метаболізм гібридного організму. Критично оцінюючи існуючі на той час теорії, М.В. Турбін (1961) справедливо констатував, що гетерозис як складне біологічне явище не може бути пояснений од-
287
ним типом взаємодії генів. Запропонована ним теорія генетичного балансу враховує причинно зумовлені зв’язки в системі ген-ознака, тобто різнонаправлену дію на гібридний організм всіх елементів спадкової основи його генотипу. Іншими словами, домінування, наддомінування, неалельні взаємодії генів є складовими генетичного балансу. Пояснення гетерозису, як наслідку неалельної взаємодії генів, що знаходяться у різних геномах (теорія гетерогеномності), знаходимо у Ю.П. Мірюти (1971).
На основі молекулярно-генетичних досліджень проблеми гібридної сили Кетчсайд і Фінч розробили теорію генетичної комплементації. Вплив гетерозису на фізіолого-біохімічні особливості організмів і його прогнозування успішно вивчали багато інших дослідників, зокрема, В.К. Шумний, В.Г. Шахбазов, Л.В. Хотильова.
Суттєвим внеском у вивчення механізмів гетерозису стало формулювання у 1983 р. В.А. Струнниковим гіпотези про компенсаційний комплекс генів (ККГ). Було чітко показано, що у відповідь на негативні мутації, які послаблюють життєздатність у популяціях мутантів, відбираються гени, котрі в гетерозиготі у F1 дають гетерозисний ефект. Пізніше ним була запропонована схема отримання ліній з високою комбінаційною здатністю.Така гіпотеза нині є панівною і досить добре пояснює вагому частку наукових фактів, добутих експериментальним шляхом, у різних біологічних об’єктів, у тому числі й у цукрових буряків. Грунтуючись на ній, Соколов В.А. (1992) для цілеспрямованого одержання і вдосконалення ліній для гетерозисної селекції розробив методику “доборів на генотипному модифікаційному фоні”. Він встановив конкретний механізм гетерозису, вважаючи, що причиною гетерозису є епістатична взаємодія генів.
Проте в літературі немає єдиної думки про ступінь впливу епістазу і його ролі у спадкуванні кількісних ознак у різних гібридів. Л.А. Тарутіна із співр. (1986) як тест на неалельні взаємодії застосувала порівняльні оцінки три- і чотирилінійних гібридів від відповідних середніх простих гібридів. Виявлено, що у більшості гібридів неалельні взаємодії не є вирішальними в генетичному контролі такої господарсько цінної ознаки, як маса зерна у качані гібридів F1 кукурудзи.
Незначимість епістатичних ефектів описують і інші дослідники англійської наукової школи, вважаючи, що епістаз може бути важливим лише в унікальних генетичних поєднаннях, але вони зустрічаються надзвичайно рідко. Проте однозначності у трактуванні генних взаємодій не було, оскільки у інших дослідах описані значущі ефекти, пов’язані з епістатичною дією генів у парних міжлінійних і трилінійних гібридів. У деяких окремих комбінаціях схрещування епістаз, як основна причина гетерозису, виявлений Спрегом і Томасом.
288
Використовуючи більш точні методи досліджень, а саме біохімічні маркери (алоферменти) для вивчення генетики кількісних ознак, Штубером і Едвардсом отримана додаткова інформація щодо різних типів дії генів. Для важливих господарсько цінних ознак найбільш важливими були і найчастіше виявлялися адитивні дії генів і наддомінування.
Незважаючи на численні спроби пояснити детермінацію гетерозису різними типами генних взаємодій, єдиної теорії, яка б могла точно передбачати результативність гібридизації, донині так і не створено. Це частково пояснюється тим, що за гібридизації батьківських форм взаємодіють не окремі гени, що зумовлюють ознаки, а цілі генні комплекси із присутніми у них зчепленням, плейотропією, генною взаємодією, кореляціями і т.д. Проте незаперечним вважається той факт, що саме гібридизація і отримання гетерозисних гібридів є резервом підвищення продуктивності. Міжсортові схрещування перевищують за продуктивністю сорти, а міжлінійні схрещування, особливо з використанням цитоплазматичної чоловічої стерильності (ЦЧС) забезпечують ще більший ефект гетерозису. Цей напрям був, є і залишається перспективним у селекційній роботі. Використання гібридів, створених на основі ЦЧС, дозволяє підняти збір цукру на 5-20 %. Слід зазначити, що сама гібридність не може бути основною причиною підвищення продуктивності. Має значення ефект від гібридизації батьківських форм, тобто цінність компонентів схрещування, селекція яких має бути побудована на генетичних принципах з використанням ефективних методів.
У практичній селекції вважається доведеним той факт, що саме міжлінійні гібриди мають перевагу перед міжсортовими і сортовими. Якщо в міжсортових або популяційних матеріалах через невирівняність вихідних форм з різною частотою зустрічаються низько- і середньоврожайні біотипи, то у міжлінійних гібридів рослини мають більший врожай у перерахунку на одну особину. Теоретичною основою цього явища є те, що різні алелі (домінантні і рецесивні) фіксовані в різних лініях, а це означає, що в поколінні F1 за гібридизації різних інбредних ліній досягається гетерозиготність локусів. Це можна умовно представити (за Бороєвичем, 1984) таким чином :
лінія із сорту А |
х |
лінія із сорту В |
♀ АВсDеGij і т.д. |
↓ |
♂ авСdЕgIJ і т.д. |
F1 AaBbCcDdEeGgIiJj і т.д.
Формула кількісного обґрунтування гетерозису Н/F1=∑ dy2, яку наводив у 1964 р. Д. Фальконер, вказувала на те, що за значення d (домінантний ефект значної кількості генів) і у2 – квадрата різниці за частотою генів між лініями, які схрещуються, різниця у2 буде най-
289