

9. Эволюция энзиматических систем, участвующих во взаимодействии клеток с кислородом.
У современных организмов 3 типа энергетического обмена: фотосинтез, дыхание, брожение. Согласно принятым представлениям материальным и энергетическим, фундаментом служит органическое соединение, возникшее из неорганического абиогенным путем. Согласно теории Апарина, первичные формы живых существ были гетеротрофы, осуществляющие свой энергообмен анаэробным путем(брожение). Современные анаэробы -гетеротрофы, имеющие активную систему энергообмена, включая дегидрогеназы, изомеразы, трансферазы, системы синтеза АТФ. Следующим этапом биоэнергетики является фототрофность, которая неразрывно связана с образованием каталитических систем, принципиально новый тип - пигменты, обладающие способностью использовать поглощенные кванты света для перехода электронов в состояние возбуждения. Преобразование энергии в химическую, достигается при участии транспортной цепи электронов. Начальным звеном в эволюции фототрофов были фототрофные бактерии, которые в качестве донора электронов используют высоковосстановительные соединения. Восстановление НАД осуществляется у бактерий в основном за счет АТФ. В дальнейшем ходе эволюции возникли организмы, обладающие механической мобилизации электронов от предельно-окислительных форм Н, в которых он находится в молекуле Н2О. Такие организмы - зеленые растения, обладающие способностью восстанавливать НАДФ, появляется свободный кислород.
10. Пути диссимиляции углеводов.
Дыхательные циклы – гликолиз и цикл ди- и трикарбоновых кислот, ПФП и прямое окисление сахаров – система взаимосвязанных процессов. Взаимосвязь гликолиза и ПФП с прямым окислением сахаров осуществляется через глюконовую кислоту и фосфотриозы.
В клетке гликолиз и ПФП пространственно не отделены друг от друга. Эти процессы протекают в растворимой части цитоплазмы, в проплостидах и в хлоропластах. Они имеют общие субстраты – гл-6-ф, фр-6-ф и 3-ФГА. В норме доля ПФП в общем дыхательном обмене составляет 10-40%. В анаэробных условиях гликолиз доминирует над ПФП. Однако в хлоропластах активность окислительного апотомического пути намного выше по сравнению с гликолизом. В цитоплазме большая часть продуктов ПФП метаболизируется через гликолиз.
Активность ПФП увеличивается при неблагоприятных условиях: засухе, калийном голодании, инфекции, затенении, засолении, при старении. Скорость окисления НАДФН (подавляющее действие продуктов одного пути дыхания на реакции другого) играет существенную роль в регуляции соотношения различных дыхательных циклов.
11. Гликолиз, суть его реакций, энергетика. Синтез сахаров при обращении гликолиза.
Реакции гликолиза идут в цитозоле и в хлоропластах. В результате гликолиза из одной молекулы глюкозы образуется 2 молекулы пировиноградной кислоты и 4 молекулы АТФ
Гликолиз – процесс анаэробного распада глюкозы, идущий с освобождением энергии, конечным продуктом которого является ПВК. Реакции гликолиза протекают в растворимой части цитоплазмы и в хлоропластах. Полностью процесс гликолиза расшифровали немецкие биохимики Эмбден, Мейергоф и советский биохимик Парнас.
гликолиз одной молекулы глюкозы м. б. выражен след. уравнением:
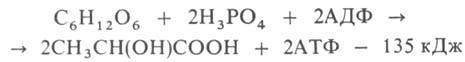
Субстратами гликолиз кроме глюкозы могут быть другие моносахариды, а также полисахариды. У растений субстратом для гликолиза может служить крахмал. Первые этапы гликолиз, спиртового и некоторых других видов брожения сходны. Стадии, в которых осуществляются необратимыми реакциями (2-4), играют существенную роль в регуляции скорости гликолиза. Наиболее важный регуляторный фермент - фосфофруктокиназа, катализирующая реакцию III; ее активность ингибируется АТФ, НАДН, лимонной и жирными кислотами, стимулируется АДФ и АМФ. Реакции II и IV катализируются соответственно гексокиназой и пируваткиназой, активность которых регулируется адениловыми нуклеотидами, промежуточными продуктами гликолиза и цикла трикарбоновых кислот. В условиях недостаточности кислорода гликолиз – единственный процесс, поставляющий энергию для осуществления физиологических функций организма. В аэробных условиях гликолиз - первая стадия окислителительного превращения углеводов: в присутствии О2 пировиноградная кислота может подвергаться дальше окислительному декарбоксилированию, а образующаяся уксусная кислота в виде СН3С(О)КоА (КоА-остаток кофермента А) полностью окисляться до СО2 и воды в цикле трикарбоновых кислот. Подавление гликолиз дыханием в присутствии О2 (эффект П а с т е р а) обеспечивает клетке наиболнн экономный механизм образования богатых энергией соединений. В тканях, где такой эффект отсутствует (напр., в эмбриональных и опухолевых), гликолиз протекает очень активно. В некоторых тканях с интенсивным гликолиз наблюдается подавление тканевого дыхания. Гликолиз - простейшая форма биологического механизма аккумулирования энергии углеводов в АТФ. Считают, что он возник в период, когда в атмосфере Земли не было О2. При энергетически более выгодном аэробном окислении из одной молекулы глюкозы образуется 38 молекул АТФ.