

2.3. Строение, свойства и биологические функции полисахаридов.
Молекулы полисахаридов включают десятки, сотни и даже тысячи моносахаридных остатков, соединенных такими же гликозидными связями, как и в составе олигосахаридов. Большинство из них образуют линейные полимеры, формирующие определённую пространственную структуру, однако некоторые полисахариды имеют разветвлённые молекулы. Моносахаридные остатки в составе полисахаридов находятся в циклической форме в виде α- или b-стереоизомеров.
Большинство полисахаридов представляют собой сложные углеводы, построенные из многократно повторяющихся остатков одного моносахарида. Однако известны полисахариды, молекулы которых состоят из остатков разных моносахаридов.
По выполняемым функциям различают запасные и структурные полисахариды. Запасные - откладываются в клетках листьев или запасающих тканей в виде упорядоченных структур - гранул. Структурные - участвуют в построении клеточных стенок растений.
Крахмал. Крахмал - основное запасное вещество растений, представляющее собой смесь двух полисахаридов - амилозы и амилопектuна, различающихся по строению молекулы и физико-химическим свойствам. Однако молекулы этих полисахаридов построены из одного моносахарида - α-D-глюкозы, находящейся в пиранозной форме.
В молекулах амилозы остатки a-D-глюкозы соединены а(1®4)-связями, образуя спиралевидно закрученные цепочечные структуры, включающие от 100 до 1-2 тыс. глюкозных остатков (рис. 1). Молекулярная масса амилозы обычно составляет от 20 до 500 тыс. Спиралевидное закручивание молекулы происходит вследствие образования водородных связей между остатками глюкозы, находящимися в соседних витках. В каждом витке амилозы содержится шесть пиранозных cтpyктyp, соединённых в цепочку гликозидными связями.
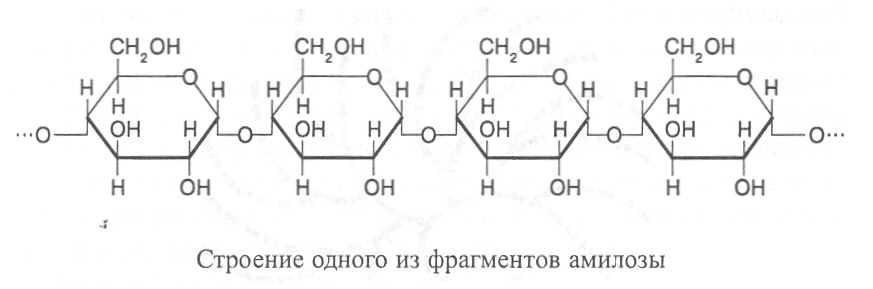
Амилоза растворяется в теплой воде и при добавлении водного раствора йода в йодистом калии окрашивается в синий цвет вследствие того, что йод образует комплeксы с остатками глюкозы. Водные растворы амилозы не отличаются высокой вязкостью и при стоянии довольно быстро образуют кристаллический осадок.
Амилопектин имеет разветвлённые молекулы, построенные из α-D-глюкозы. В точках ветвления гликозидные связи образуются между первым и шестым углеродными атомами глюкозных остатков (α(I®6)-связи). Между точками ветвления глюкозные остатки так же, как в амилозе, соединены α(I®4)-связями.
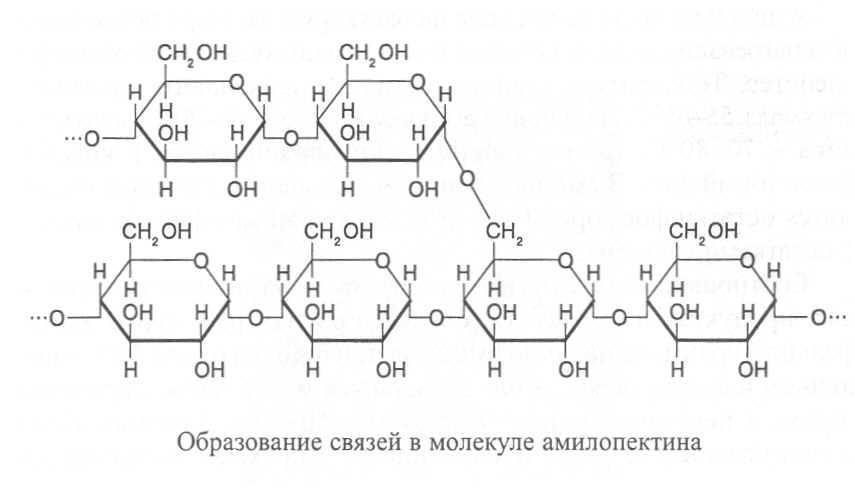
Точки ветвления в молекулах амилопектина имеются через каждые 12-15 остатков глюкозы. Молекулярная масса амилопектина значительно больше, чем у амилозы, и может достигать 1 млн. Схема строения молекулы амилопектина показана на рисунке 2.
Амилопектин в тёплой воде не растворяется, а при более сильном нагревании с водой образует очень вязкий коллоидный раствор - клейстер. Температура клейстеризации картофельного и ржаного крахмала 55-65°С, пшеничного и кукурузного - 60-70°С, крахмала риса - 70-80°C. Йодом амилопектин окрашивается в красно-фиолетовый цвет. В амилопектине в небольшом количестве содержатся остатки фосфорной кислоты, соединённые эфирной связью с остатками глюкозы.
Соотношение амилозы и амилопектина в различных растительных продуктах изменяется в очень широких пределах. В картофельном крахмале на долю амилозы приходится около 20%, пшеничном и кукурузном - около 25%, рисовом - 15-20%, в крахмале гороха и некоторых сортов кукурузы - 50-80%. Крахмал яблок почти полностью состоит из амилозы, а крахмал восковидных сортов кукурузы - только из амилопектина.
У одного и того же вида растений содержание амилозы и амилопектина в крахмале может изменяться в зависимости от фазы развития и условий внешней среды. В разных органах растений синтезируется крахмал совершенно определенного состава. Так, например, в крахмале клубней картофеля обычно содержится 19-22% амилозы, а в молодых побегах в два раза больше.
В растениях крахмал образуется в листьях как продукт фотосинтеза, а также в зерновках и семенах, клубнях, корневищах, утолщенных частях стеблей как запасное вещество. Фотосинтетический крахмал откладывается в хлоропластах в виде гранул, называемых крахмальными зёрнами, и довольно быстро используется в процессе дыхания и для синтеза других веществ. Значительная его часть превращается в транспортную форму углеводов - сахарозу, которая по флоемной системе поступает в нефотосинтезирующие органы, распадается там до глюкозы и фруктозы и в виде моносахаридов включается в различные биосинтетические процессы.
Запасной крахмал также откладывается в виде зёрен и у целого ряда растений накапливается в значительном количестве в запасающих тканях и органах. В зерне злаковых его содержание обычно составляет 50-70%, в рисе -75-80%, в зерне зернобобовых культур - 30-50%, в клубнях картофеля - 12-20%, в клубнях батата, ямса и маниока - 20-30%, в листьях растений - до 1-2%.
Крахмальные зёрна чаще всего имеют вид овальных или сферических частиц (рис. 3), имеющих разную форму и размеры (2-170 мкм). Под микроскопом можно различить их слоистое строение. Размеры и строение крахмальных зёрен у разных видов и даже сортов растений имеют характерную специфику и могут использоваться для идентификации генотипов, а также обнаружения примесей одного растительного продукта в другом.
Запасной крахмал вначале откладывается в пластидах, называемых амилопластами. По мере наполнения происходит постепенная деградация их мембранной структуры и они превращаются в крахмальные зерна.
Крахмалоносные растения представляют легковозобновляемое сырье для перерабатывающей промышленности, которое используется для получения продовольственного и технического крахмала, глюкозы, этилового спирта и даже пластмасс, обладающих высокой прочностью и экологической безопасностью (при сгорании не дают ядовитых выделений).
Полифруктозиды. В растениях семейств лилейные, мятликовые, астровые, колокольчиковые синтезируются запасные углеводы, построенные из 4-40 остатков b-D-фруктозы, в связи с чем их называют полифруктозидами, или фруктанами. Остатки фруктозы в их молекулах соединены гликозидными связями, образующимися между вторым и первым углеродными атомами (b (1®2)-связи).
Полифрутозиды содержатся в листьях, корнях, семенах указанных выше растений, накапливаются в значительном количестве в нижней утолщённой части стеблей мятликовых трав (до 6-8% сухой массы) и в созревающих зерновках злаковых культур (рожь, пшеница, ячмень, овёс). В листьях они являются основными продуктами фотосинтеза, тогда как фотосинтетический крахмал у этих растений не образуется.
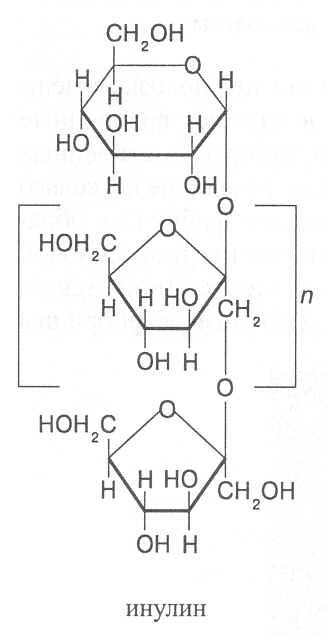
Из полифруктозидов наиболее хорошо изучен инулин, содержащий в молекуле 37-44 фруктозных остатка. К одному из концов молекулы инулина присоединён остаток α-D-глюкозы. Молекулярная масса инулина 5-6 тыс. Он хорошо растворяется в горячей воде, не обладает восстановительными свойствами, хорошо усваивается организмами человека и животных, в связи с чем растения, способные накапливать инулин, используются как кормовые культуры и как сырьё для промышленного получения фруктозы. Фруктозу получают из инулина путём его кислотного гидролиза.
Большое количество инулина содержится в клубнях георгина и артишока (до 50%), топинамбура (10-12%), корнях цикория (свыше 10%). В чесноке общее содержание полифруктозидов достигает 20-30% и половину из них составляет инулин.
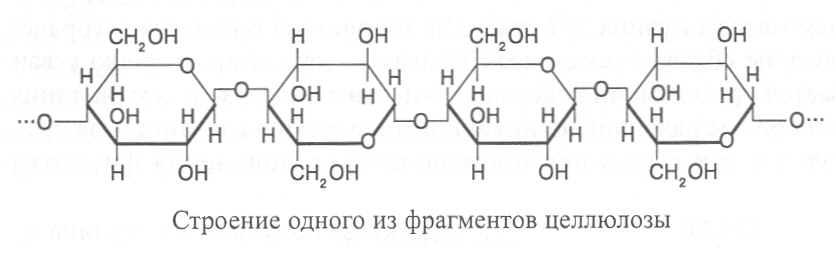
Целлюлоза. Целлюлоза, или клетчатка - довольно устойчивое вещество волокнистого строения, не растворяется в воде и органических растворителях, однако хорошо растворимо в аммиачном растворе гидроксида меди (реактив Швейцера). Молекулы целлюлозы состоят из остатков b-D-глюкозы, соединенных b(1®4)-связями. В каждой молекуле целлюлозы может содержаться 1500-10000 пиранозных остатков b-D-глюкозы, образующих неразветвлённый полимер.
Между линейно вытянутыми молекулами целлюлозы, имеющими свободные гидроксильные группы, возникают водородные связи, с помощью которых нитевидные полимеры, построенные из остатков глюкозы, объединяются в пучки, включающие несколько десятков молекул. Такие целлюлозные пучки, или фибриллы, обладают очень высокой прочностью и служат структурной основой клеточных стенок растений. Как видно на электронной микрофотографии клеточной оболочки (рис. 4), целлюлозные фибриллы размещаются слоями, образуя сетчатую структуру, сквозь которую свободно проникает вода с растворенными в ней веществами.
Целлюлоза в том или ином количестве содержится во всех растительных тканях. Особенно много целлюлозы в растительных волокнах (хлопковом, льняном) - 80-95%, древесине и соломе - 40-50%. В других растительных продуктах её значительно меньше: зерно злаковых и зернобобовых культур – 2-6%, зерно пленчатых злаков - 7-14%, семена масличных - 5-25%, клубни картофеля - около 1 %, корнеплоды - 0,5-1,5%, овощи 0,5-1,2% (томаты - 0,2%), плоды и ягоды - 0,5-2%, вегетативная масса кормовых трав - 20-30% (последний показатель - в расчёте на сухую массу).
Целлюлоза практически не усваивается организмами человека и нежвачных животных, тогда как жвачные животные способны её усваивать с помощью ферментов микроорганизмов, обитающих в преджелудках этих животных и участвующих в процессах пищеварения.
При нагревании с раствором кислоты целлюлоза подвергается гидролизу, превращаясь в глюкозу, которая используется как источник углерода для культивирования дрожжевых клеток с целью промышленного получения этилового спирта и кормовых дрожжей с повышенным содержанием белков и витаминов. При этом в качестве источника целлюлозы служат отходы древесины и целлюлозосодержащие растительные остатки - солома, корзинки подсолнечника, льняная костра, стержни кукурузных початков, свекловичная меласса, картофельная мезга, хлопковая шелуха и др. Большое количество целлюлозы расходуется для химической переработки.
В построении клеточных стенок растений наряду с целлюлозой участвуют также другие структурные полисахариды - гемицеллюлозы и пектиновые вещества, которые связаны с молекулами целлюлозы водородными связями.
Гемицеллюлозы. Гемицеллюлозы - это смесь полисахаридов, образу-ющих при гидролизе маннозу, галактозу, ксилозу, арабинозу и уроновые кислоты - глюкуроновую и галактуроновую. Они нерастворимы в воде, но растворяются в щелочных растворах. В клеточных стенках растений содержание гемицеллюлоз составляет около 30%. Много их накапливается в древесине и соломе (10-30%), оболочках семян, кукурузных початках, отрубях, вегетативной массе растений. Разные виды растений заметно различаются по составу гемицеллюлоз.
ГАЛАКТАНЫ. Их молекулы построены из остатков b-D-галактозы, соединённых b(1®4)-связями. В каждой молекуле объединяются более 100 остатков галактозы.

Галактаны содержатся в составе клеточных стенок многих растений, особенно много их в семенах люпина.
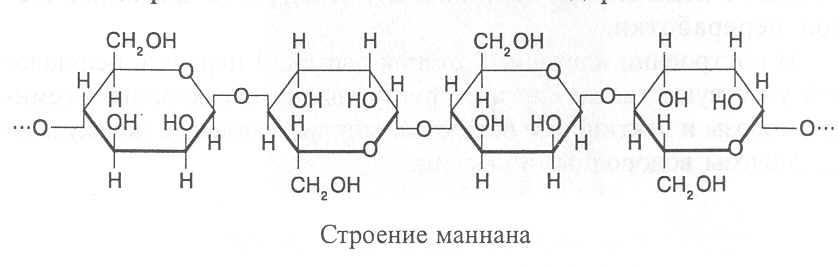
МАННАНЫ. Остатки маннозы в маннанах соединены b(1®4)-связями. В каждой молекуле насчитывается от 200 до 400 моносахаридных единиц. Много маннанов содержится в древесине хвойных деревьев и в клеточных стенках водорослей.
КСИЛАНЫ. Их молекулы построены из остатков b-D-ксилозы в пиранозной форме, соединённых b(1®4)-связями. В составе полимера могут находиться до 200 ксилозных остатков. В соломе и древесине содержание ксиланов достигает 25-28%.
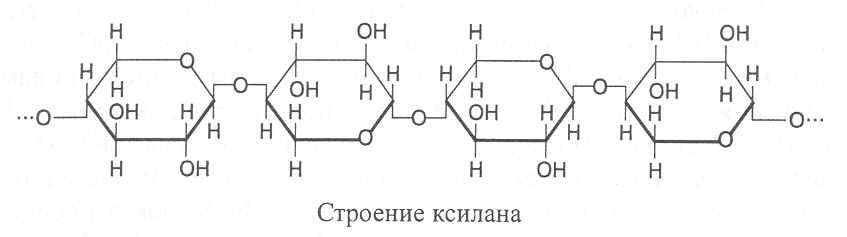
В молекулах ксиланов обычно имеются ответвления в виде остатков арабинозы, а также глюкуроновой и галактуроновой кислот. Ответвления чаще всего образуются за счёт этерификации третьего углеродного атома ксилозы. Карбоксильные группы остатков уроновых кислот образуют эфиры с метиловым спиртом. Ксиланы разных растений отличаются частотой и моносахаридным набором ответвлений в молекуле.
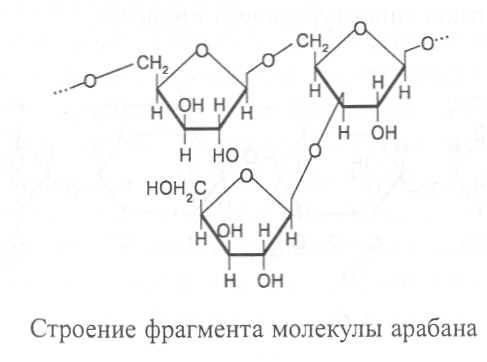
АРАБАНЫ. Это полисахариды клеточной стенки растений, которые состоят из остатков a-L-арабинозы, соединённых гликозидной связью между первым и пятым углеродными атомами. При этом к каждому второму остатку арабинозы в линейной структуре присоединён в виде ответвления еще один остаток арабинозы. В ответвлениях связь образуется между третьим углеродным атомом арабинозы, находящейся в цепочке, и первым углеродным атомом бокового остатка арабинозы.
Как и целлюлоза, гемицеллюлозы не усваиваются организмом человека, но могут усваиваться жвачными животными с помощью ферментов микроорганизмов, находящихся в преджелудках.
ГЛЮКАНЫ. К глюканам относятся полисахариды, образуемые из b-D-глюкозы, но в их молекулах остатки глюкозы соединяются не только b(1®4) -связями, как в целлюлозе, но также и b(1®3)-связями или только b(1®3)- связями. К таким полисахаридам относятся каллоза и лихенин. Каллоза - полисахарид, включающий до 100 остатков b-D-глюкозы в молекуле, соединённых b(1®3)-связями. Она содержится в ситовидных трубках флоэмной системы растений. В молекулах лихенина остатки b-D-глюкозы соединены как b(1®4)-связями, так и b(1®3)-связями (встречаются с частотой около 30%). Лихенин входит в состав клеточных стенок растений, особенно много его в лишайниках.
Пектиновые вещества. Пектиновые вещества в растениях представлены двумя группами соединений - пектинами и протопектинами, которые различаются строением и физико-химическими свойствами.
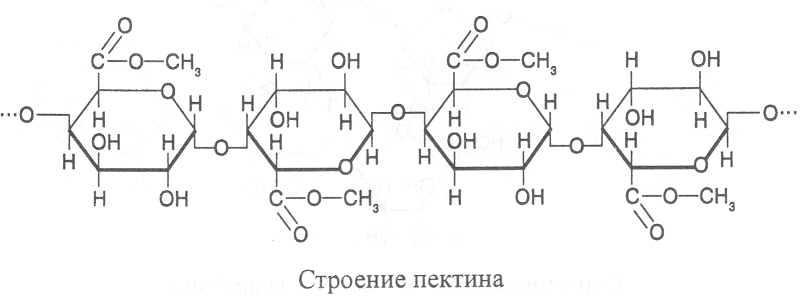
Пектины - водорастворимые полисахариды, построенные из остатков α-D-галактуроновой кислоты, которые соединены α(1®4)-связями. Большая часть карбоксильных групп остатков галактуроновой кислоты связана эфирными связями с остатками метилового спирта, а к другим карбоксильным группам присоединены катионы кальция или магния. В каждой молекуле пектина содержится более 100 остатков галактуроновой кислоты.
Основная масса пектиновых веществ растений представлена протопектином, который находится в структуре клеточных стенок. Протопектин образуется в результате связывания эфирными связями пектина с галактанами и арабанами, входящими в состав клеточной стенки растения. Эфирные связи возникают между карбоксильными группами пектина и гидроксильными группами гемицеллюлоз.
Полисахариды протопектина нерастворимы в воде и имеют более высокую молекулярную массу по сравнению с пектинами. Много протопектина накапливается в формирующихся плодах rpуши, яблони, цитрусовых, айвы, что обусловливает их жёсткую консистенцию. При созревании плодов происходит превращение протопектинов в пектины, вследствие чего их консистенция становится мягкой.
Общее содержание пектиновых веществ в плодах и ягодах составляет 0,3-1,5%, в корнеплодах - 1,5-2,5%, клубнях картофеля 0,1-0,5%, в томатах - 0,1-0,2%, в капусте - 0,3-2,0%, в кожуре апельсина и лимона - 4-7%.
Характерная особенность пектиновых веществ плодов и ягод - способность образовывать желе, или студни, в насыщенном растворе сахара (65-70%) и кислой среде (рН 3,1-3,5). Лучшей желирующей способностью обладают более высокомолекулярные полисахариды пектиновых веществ.
В стеблях льна пектиновые вещества скрепляют между собой волокна. Для отделения раcтитeльных волокон производится росяная или водяная мочка льносоломы, при которой происходит гидролиз пектиновых веществ под действием ферментов микроорганизмов.
Камеди и слизи. Это растворимые в воде полисахариды, образующие очень вязкие растворы вследствие их набухания.
Растительные камеди выделяются на стволах и ветвях некоторых деревьев (вишневых, сливовых, миндальных) в виде клейких наплывов при повреждениях. При гидролизе они дают галактозу, маннозу, рамнозу, арабинозу, ксилозу, а также уроновые кислоты.
Слизи, откладываясь между плазмалеммой и клеточной стенкой, способствуют удерживанию воды в клетках и полостях растения, защищают от проникновения инфекции. При их гидролизе в основном образуются пентозы (арабиноза и ксилоза), а также небольшое количество галактозы, глюкозы и фруктозы.
Много слизей содержится в семенах льна, клевера, люцерны, ржи и некоторых других растений. Повышенная вязкость ржи при размоле вызвана наличием именно слизей, вследствие чего зерно ржи размалывается значительно труднее, чем пшеница. Содержащиеся в ржаной муке слизи замедляют гидролитические процессы при формировании теста и тем самым улучшают его формоудерживающую способность.
Камеди и слизи из различных растительных источников существенно отличаются набором и удельным соотношением образующих их полисахаридов. Они состоят из молекул разной степени полимеризации, многие из которых имеют довольно высокую степень ветвления.
Вопросы для повторения.
1. Каковы структурные особенности стереоизомеров моносахаридов, относящихся к D- или L-ряду? 2. Как образуются циклические формы моносахаридов и в чём состоят различияа- иb-стереоизомеров? 3. Как записывается структура пиранозных и фуранозных форм моносахаридов с помощью формул Хеуорса? 4. Какие образуются конформации молекул у гексоз и пентоз? 5. Как образуются окисленные и восстановленные производные, а также фосфорнокислые эфиры моносахаридов? 6. В чём состоят особенности образования гликозидов, дезокси- и аминопроизводных моносахаридов? 7. Каковы структурные и биологические особенности важнейших альдоз и кетоз? 8. Как образуются молекулы сахарозы, мальтозы, целлобиозы,b-левулина и других олигосахаридов? 9. Из каких моносахаридов и по какому принципу строятся молекулы важнейших полисахаридов - крахмала, полифруктозидов, целлюлозы и гемицеллюлоз, пектиновых веществ, камедей и слизей? 10. Какие биологические функции выполняют указанные выше олигосахариды и полисахариды? 11. Каково содержание сахаров и различных полисахаридов в растительных продуктах? 12. Какие моносахариды и олигосахариды относятся к редуцирующим сахарам? 13. Какое значение имеют углеводы в формировании качества растительных продуктов? 14. Из каких основных компонентов состоит крахмал и каковы строение и свойства этих компонентов? 15. Какие известны разновидности гемицеллюлоз и пектиновых веществ?
Резюме по модульной единице 2.
Углеводы являются важными компонентами клеток живых организмов. Одни из них служат основным дыхательным материалом организмов (сахара, крахмал, по-лифруктозиды) и откладываются в качестве запасных веществ, другие выполняют структурные (целлюлоза, гемицеллюлозы, протопектин) и защитные (камеди и слизи) функции. Большинство сахаров существуют в форме стереоизомеров и относятся, как правило к D-ряду. Гексозы и пентозы существют в организмах преимущественно в виде циклических форм (пиранозных или фуранозных).
Из моносахаридов образуются восстановленные (многоатомные спирты), окисленные (альдоновые, альдаровые и уроновые кислоты) производные, фосфорнокислые эфиры, амино- и дезоксипроизводные, гликозиды, которые являются продуктами превращения моносахаридов и участвуют в метаболизме. Легкоусвояемые формы углеводов (сахара, крахмал, полифруктозиды, пектиновые вещества) способны накапливаться в значительном количестве в растительных продуктах и поэтому определяют их питательную и техническую ценность.
Олигосахариды образуются из моносахаридов, остатки которых соединяются в молекулах олигосахаридов О-гликозидными связями. Сахароза и олигофрукто-зиды откладываются в запасающих органах растений или используются в качестве транспортных форм углеводов в растительных организмах. Мальтоза – продукт распада крахмала, целлобиоза в свободном виде не накапливается, так как используется в синтезе целлюлозы.
Молекулы полисахаридов построены из остатков моносахаридов, которые соединяются, как и в молекулах олигосахаридов, О-гликозидными связями. Молекулы амилозы, полифруктозидов, пектинов, маннанов, галактанов представляют собой цепочечные полимеры. Молекулы амилопектина, ксиланов, арабанов, камедей и слизей имеют ответвления разной степени сложности. Крахмал, полифруктозиды, пектины откладываются в запасающих органах растений. Целлюлоза, гемицеллюлозы, протопектин участвуют в построении клеточных стенок растений.
Модульная единица 3. Липиды.
Цели и задачи изучения модульной единицы. Изучить строение, свойства и биологические функции основных групп липидов. Научить студентов использовать сведения о липидах при оценке качества растительной продукции.
К липидам относятся вещества, различающиеся по химическому составу, строению и выполняемым функциям, но обладающие близкими фифизико-химическими свойствами. Все они содержат гидрофобные радикалы и группировки, вследствие чего не растворяются в воде, но хорошо растворимы в неполярных органических растворителях - эфире, бензине, бензоле, хлороформе.
В зависимости от химического состава и строения липиды под-разделяют на три класса: простые, сложные и стероидные.
Простые липиды представляют собой сложные эфиры спиртов и вы-сокомолекулярных карбоновых кислот, к ним относятся жиры и воски. Сложные липиды содержат в составе молекул, кроме спирта глицерина и карбоновых кислот, остатки других соединений: ортофосфорной кислоты, азотистых оснований, моносахаридов и др. Они образуют две группы веществ - фосфолипиды и гликолипиды.
Стероидные липиды - это циклические соединения, являющиеся про-
изводными циклопентанопергидрофенантрена и представленные как сво-
бодными стероидными веществами, так и связанными формами в виде гли-
гликозидов и эфиров. К липидам очень часто относят также растворимые в жирах витамины и пигменты.
Общее содержание структурных липидов в вегетативных частях растений находится в пределах 0,1-0,5%. Накопление запасных липидов в семенах различных растений достигает следующих величин: зерно злаковых и зернобобовых культур - 1-8%, соя и хлопчатник - 20-30%, подсолнечник, арахис, лен, конопля, рапс, горчица, маслины - 20-50%, мак, клещевина, ядра орехов - 50-60%, в зародышах зерновок пшеницы - 8-14%, кукурузы - 30-40%. Растения с высоким содержанием в семенах запасных липидов выделены в особую группу масличных культур. Известны также растения - накопители воска.