

Раздел I. Аппарат пищеварения
(Apparatusdigestorius)
В онтогенезе хордовых система органов пищеварения формируется преимущественно из двух зародышевых листков: мезодермы и энтодермы; только выстилка самой передней и самой задней её частей является производной эктодермы. Пищеварительная система анатомически делится на четыре отдела: головная кишка (ротовая полость и глотка), передняя кишка (пищевод и желудок), средняя кишка (тонкий кишечник и связанные с ним застенные железы) и задняя кишка (толстый кишечник). (Клоака, через которую происходит выброс непереваренных частиц пищи у представителей многих групп позвоночных, рассматривается как часть мочеполовой системы.)
Аппарат пищеварения в целом выполняет четыре функции:
захват пищи;
её механическое измельчение и перемешивание и перемещение (эвакуация) по пищеварительному тракту;
переваривание (химическая обработка – разложение высокомолекулярных соединений до низкомолекулярных, какие могут быть перенесены во внутреннюю среду организма);
выброс непереваренных остатков (формирование, дегидратация (высушивание) каловых масс и выталкивание их).
ГОЛОВНАЯ КИШКА
Ротовая полость(cavumoris) открывается наружу ротовым отверстием, в её составе у животных разных классов имеются в различной степени развитые губы, язык, зубы, нёбо, слюнные железы и другие органы.Глоткой(farinx) называют часть кишечной трубки, где осуществляется анатомическая связь пищеварительной и дыхательной систем: либо в её стенках открываются жаберные щели, либо через глотку от хоан до гортани проходит воздух. Глотка может быть резко отделена от ротовой полости, а может не иметь заметной границы, тогда говорят оротоглоточной полости(cavum orofaringaeus).
Строение ротоглотки у ланцетника, при всём своеобразии, чётко демонстрирует его принадлежность к типу Хордовые: боковые стенки глотки пронизаны многочисленными (до 120 с каждой стороны)жаберными щелями. В остальном ротоглотка ланцетника скорее несёт черты, унаследованные от «нехордовых» предков - фильтраторов.
Перед входом в ротоглотку у ланцетника имеется довольно широкая, окружённая щупальцами предротовая воронка. Маленькое ротовое отверстие находится на её дне и окружено тонкой складкой –парусом. Ротовое отверстие ведёт непосредственно в широкую и длинную глотку. На дне глотки проходитподжаберная борозда(эндостиль), выстланная реснитчатым эпителием с многочисленными слизистыми клетками. Реснички гонят слизь с налипшими частичками пищи в оральном направлении, там пополукольцевым бороздамона поднимается вверх, внаджаберную борозду(устроенную так же, как поджаберная). Наджаберная борозда ведёт в кишку, у начала которой глотка резко сужается. Профильтрованная вода удаляется через жаберные щели ватриальную полость, а оттуда, черезатриопор– наружу.
Ротовое отверстие у круглоротыхоформлено хрящамиротовой (присасывательной) воронки, которые выполняют функцию отсутствующих челюстей. Слизистая оболочка ротовой воронки несёт роговые «зубы» (у миног). Из воронки отверстие ведёт в собственно ротовую полость. Строение ротовой полости прекрасно приспособлено к способу питания миног и миксин – паразитов и падальщиков.Языккруглоротых не гомологичен языку остальных позвоночных, хотя и является мощным мышечным органом и по функциональным особенностям напоминает язык млекопитающих. При движении назад язык помогает присасываться к жертве (при движении языка вперёд миноги и миксины удерживаются на теле жертвы роговыми «зубами» ротовой воронки). На поверхности языка также имеются ряды роговых зубчиков, которыми круглоротые, как тёркой, скоблят ткани жертвы. В основании языка имеютсяпристенные слюнные железы, используемые для наружного пищеварения.Глоткакак место анатомической связи пищеварительной и дыхательной систем у взрослых круглоротых фактически отсутствует (имеется только в личиночной стадии). Из ротовой полости ведут два отверстия: вверх – впищеводи вниз – вдыхательную трубку, на границе отверстий имеется складка слизистой оболочки –парус. У свободноплавающей миноги или миксины парус закрывает отверстие пищевода, а у присосавшейся – отверстие дыхательной трубки. (Механизм дыхания круглоротых будет рассмотрен в соответствующей главе.)

У хрящевых рыбпоявляются
настоящие челюсти, сформированные
нёбно-квадратными и Меккелевыми хрящами.
На коже, покрывающей челюсти, плакоидные
чешуи преобразовались в крупные
плоско-конические (в большинстве случаев)зубы(dentes) в несколько рядов. Такие
же зубы развиваются укостных рыбна челюстях, образованных первичными
и вторичными костями, и на нёбе. У рыб
встречаются и плоские давящие зубы
(например, у моллюскоядных). По расположению
на челюсти зубы акродонтные, по сменности
полифиодонтные. Многие рыбы имеют ещё
и так называемые глоточные зубы, которые
располагаются на нижних члениках
последней жаберной дуги. Это давящие
зубы с плоской
 поверхностью,
напротив них в крыше глотки лежит плоский
роговойжерновок, опирающийся на
кости осевого черепа.
поверхностью,
напротив них в крыше глотки лежит плоский
роговойжерновок, опирающийся на
кости осевого черепа.
Язык(lingua) развит слабо, по сути дела, это просто складка слизистой оболочки дна ротовой полости, поддерживаемая копулами подъязычной и жаберных дуг. Собственной мускулатуры язык не имеет. Исключением является брызгун, способный языком, как поршнем, с силой выталкивать воду из ротовой полости, сбивая насекомых на лету. В эпителии слизистой оболочки ротовой полости рыб имеются отдельныеодноклеточные слизистые железы.
Заметной границы ротовой полости с глоткой у рыб нет. В боковых стенках глотки располагаются жаберные щели, каудально глотка резко сужается и переходит в пищевод.
Зубы позвоночных классифицируют:
1) по положению на челюсти
– акродонтные (размещаются на краю челюсти);
– плевродонтные (размещаются на медиальной поверхности края челюсти);
– текодонтные (размещаются в альвеолах);
2) по сменности
– полифиодонтные (многосменные);
– дифиодонтные (двусменные);
– монофиодонтные (односменные);
3) по форме
– конические;
– листовидные;
– плоские;
Зубы млекопитающих, кроме того, классифицируют:
по длине коронки – брахиодонтные (короткокоронковые) и гипсодонтные (длиннокоронковые);
по форме поверхности – секодонтные (с режущей поверхностью), лофодонтные (складчатые), селенодонтные (лунчатые), бунодонтные (бугорчатые) и др.
Зубная система млекопитающих – гетеродонтная (на зубной аркаде различаются резцы, клыки, предкоренные (премоляры) и коренные (моляры) зубы); у остальных позвоночных зубная система гомодонтная – зубы одинаковые.
В строении ротоглотки амфибий имеются значительные отличия от строения ротоглотки рыб. Орально в крышу ротовой полости открываютсяхоаны(choanae) – внутренние ноздри, а каудовентрально находится отверстиегортани, ведущей в трахею. Таким образом, воздух при дыхании проходит по всей ротовой полости, при этом в ней не должно быть пищи (воды). Над гортанью располагается широкое отверстие пищевода. Дорсолатерально в задней части ротоглотки расположены отверстияглоточно-барабанных (Евстахиевых) труб, ведущих в барабанную полость. Стенки ротовой полости амфибий выполняют функцию газообмена, особенно актуальную для форм, не имеющих лёгких. В связи с этим в стенках ротовой полости находятся пучковидные разветвления кровеносных капилляров.
Языку амфибий мускулистый, у бесхвостых амфибий он негомологичен языку других позвоночных. Язык лягушки прикрепляется к переднему краю нижней челюсти, задний конец языка – свободный. Язык, сильно удлиняясь за счёт наполнения его лимфой, может «выстреливать» вперёд, захватывать добычу и забрасывать её в ротовую полость. В проталкивании пищи в пищевод – глотании – у бесхвостых амфибий язык не задействован, в этом принимают участие глазные яблоки, которые втягиваются, нажимая на пищевой объект сверху. У хвостатых амфибий, в отличие от бесхвостых, язык имеет обычное строение: его передний конец свободен, а задний связан с подъязычным аппаратом.
В ротовой полости амфибий имеются многоклеточные пристенные слизистые слюнные железы, особенно хорошо развитые у наземных форм. У взрослых амфибий некоторых видов в слюне имеется небольшое количество пищеварительных ферментов. У лягушек слюна, вырабатываемая язычными железами, клейкая, что способствует захвату пищевых объектов языком.

Зубыамфибий мелкие конические, акродонтные, полифиодонтные. Располагаются они на зубных, верхнечелюстных и межчелюстных костях, на элементах нёба. У амфибий большинства видов зубная система развита слабо, у некоторых видов зубы и вовсе могут отсутствовать (например, у жаб).
Дно ротовой полостиамфибий широкое, мускулистое, играет основную роль в присущем амфибиям гулярном дыхании (см. раздел «Дыхательная система). Вкрыше ротовой полостиамфибий наблюдаются скопления лимфоидной ткани. Их количество и расположение непостоянно, поэтому они не могут называться миндалинами, но сходство с последними очевидно.
В связи с передним расположениемхоан и отсутствием вторичного твёрдого нёба, урептилийглотка, как и у амфибий, не отделена от ротовой полости (кроме крокодилов). Нанёбе(palatum) рептилий имеется широкаянёбная щель(между нёбными пластинками). В самой передней части нёбной щели имеются парные отверстиявомеро-назального (Якобсонова) органа, непосредственно за ними – хоаны.
Каудально в крыше глотки имеются парные отверстия глоточно-барабанных (Евстахиевых) труб.
На дне ротовой полостирасположенязык. Он может иметь разную форму и степень подвижности в связи с типом питания рептилии, но, в любом случае, это мышечный орган. Язык может быть коротким, треугольной формы (некоторые ящерицы), массивным и малоподвижным (черепахи), длинным и раздвоенным (змеи). Кончик языка используется ящерицами и змеями для улавливания ароматических веществ из воздуха и доставки их к Якобсонову органу.

Гортанная щельбольшинства рептилий сильно выдвинута вперёд и хорошо видна на спинке языка, а широкое отверстие пищевода открывается значительно каудальнее. Это облегчает рептилиям дыхание при проглатывании крупной добычи.
Зубная системау рептилий гомодонтная. Зубы простые конические, у ящериц и змей чаще плевродонтные, но бывают и акродонтные; у крокодилов – текодонтные. Акродонтные зубы – монофиодонтные, другие – полифиодонтные. Ядовитые зубы змей имеют акродонтное прикрепление с помощью связок и при закрывании рта «складываются» внутрь ротовой полости. Такие зубы имеют либо канал, либо желобок на внутренней поверхности, по которому стекает секрет ядовитых желёз. Черепахи лишены зубов, вместо этого их челюсти одеты роговой рамфотекой.
Практически во всех мягких поверхностях ротоглотки рептилий имеются многоклеточные пристенные слюнные железы. Белковый компонент содержится только в секрете ядовитых желёз (по происхождению они тоже слюнные).
Значительные отличия имеются в строении ротоглотки крокодила. В связи с появлением вторичного твёрдого нёбахоаны у крокодилов оказываются отодвинуты назад и открываются непосредственно над гортанной щелью. Прикрепление зубов у крокодилатекодонтное. Такое прикрепление обеспечивает значительно большую прочность зубам при удержании крупной добычи. Язык крокодила слабо отделён от дна ротовой полости.
Для ротоглотки птицхарактерно отсутствие вторичного неба и расположениехоанв глубокой срединной щели, что способствует некоторому разобщению дыхательных и пищеварительных путей. Губы заменены роговым чехлом клюва –рамфотекой, которая постоянно нарастает и либо постепенно стирается и слущивается (куры, попугаи), либо претерпевает быструю смену со слущиванием старого слоя в период сезонной линьки. У большинства птиц вся рамфотека имеет постоянный радиус кривизны, что позволяет ей равномерно расти и равномерно изнашиваться. У птиц, клюв которых имеет несколько разных радиусов кривизны, позади рамфотеки находится участок верхней челюсти, покрытый кожей – восковица (у соколообразных), или же рамфотека состоит из нескольких пластинок, нарастающих независимо (у буревестникооюразных, многих пеликанообразных).
Зубов у современных птиц нет; нет и преддверия ротовой полости. Функцию зубов выполняет рамфотека. Она может иметь острые режущие края для разрезания мяса или вылущивания семян из кожуры, крючки или зубцы для удержания скользкой добычи, пластинки для отцеживания планктона.
Языку птиц, в том случае, если он нормально развит, повторяет форму дна ротовой полости. Его подвижность, даже такая высокая, как у дятлов, определяется подвижностью подъязычной кости, так как собственных мышц язык не имеет (исключение – попугаи). Язык используется в основном для манипулирования пищевыми объектами в ротовой полости, но может иметь отношение и к их захвату. В связи с последним, он имеет трубкообразную форму у колибри, снабжен щеточкой из нитевидных сосочков также для сбора нектара у попугаев лори, или пластинками для отцеживания планктона у уток и фламинго. Вместе с тем, есть птицы, у которых язык сильно уменьшен (значительно короче клюва) или отсутствует – бескилевые, буревестникообразные, большинство пеликанообразных, аисты, удодообразные.
Слюнные железыу птиц имеются толькопристенные, находящиеся на верхней челюсти, в межчелюстном пространстве, на языке и в глотке. Развиты они довольно слабо, так как существенной обработки пищи в ротовой полости у птиц не происходит, а могут и вовсе отсутствовать (пеликанообразные). Заметным исключением являются стрижи, имеющие крупные застенные слюнные железы. Выделяемый ими чрезвычайно обильный и богатый белком секрет используется при строительстве гнезда.
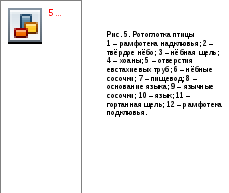
В ротовой полости у птиц имеются механические сосочки– выросты слизистой оболочки, покрытые ороговевающим эпителием. Они чаще всего собраны в 3 группы: небные, глоточные и язычные, проходящие поперечными рядами. Они ориентированы назад и способствуют продвижению пищевого объекта в сторону пищевода. Небные поверхности птиц, вылущивающих семена из оболочки, также ороговевают и имеют валики для удерживания семян в ротовой полости в правильном положении.
Отверстия евстахиевых трубрасположены в срединной щели, находящейся каудальнее щели хоан.
У многих воробьинообразных (вьюрковые, врановые) в межчелюстном пространстве находится растягиваемый мешок, используемый для временного складирования корма. У пеликанов аналогичный мешок развит еще сильнее и служит сачком для ловли рыбы, расширяющимся при раскрывании нижней челюсти.
Ротоглотка млекопитающих, в первую очередь, характерна наличием мягких и мускулистых губ, щёк и языка, используемых младенцами для сосания молока. Отверстия хоан за счёт формирования вторичного твёрдого нёба отнесены за пределы ротовой полости, в хорошо оформленную глотку, что дало возможность млекопитающим длительное время обрабатывать пищу во рту перед проглатыванием.
Губы(labia) покрыты снаружи кожей, а изнутри – слизистой оболочкой. Они снабжены огромным количествомнервных окончаний, многие из которых связаны со специфическими осязательными волосками или сконцентрированы в утолщениях на поверхности губ. Размер ротовой щели у млекопитающих разных видов связан с типом питания: углы рта - губныекомиссуры- у хищников находятся на уровне средних коренных зубов, а у травоядных копытных – перед коренными зубами. У крупных жвачных верхняя губа входит в составносогубного зеркальца– участка лишённой волос и снабжённой серозными железами кожи, простирающегося от края губы и заполняющего всю область ноздрей. У свиней на этом месте формируетсяхоботковое зеркальце.
В основе губ лежит круговая мышца рта, в которую вплетаются мышцы – дилататоры ротового отверстия. В толще слизистой оболочки, а у животных многих видов и в толще мышц, залегают пристенные слюнные железы, особенно многочисленные в области углов рта.
Щёки(bucca) имеют в своей основе щёчную мышцу. Слизистая оболочка щёк может нести ороговевающие механические конусовидные сосочки. Размер щёк связан с размером ротовой щели: чем каудальнее угол рта, тем короче оказывается щека. У травоядных животных щеки несут большую функциональную нагрузку, чем у хищных, возвращая корм на жевательную поверхность зубов при длительном пережевывании. Щёки могут быть сильно растяжимы и формироватьзащёчные мешки, которые при наполнении покрывают даже шею (хомяки). Такие мешки используются как ёмкости для хранения и переноса пищи. Пространство между щеками и губами с одной стороны и зубами с другой называетсяпреддверием ротовой полости.
В толще щёк залегают щёчные слюнные железы, особенно хорошо развитые у травоядных.
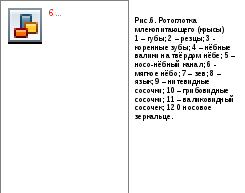
Нёбомлекопитающих длинное, состоит из двух частей: твёрдого и мягкого нёба. Основойтвёрдого нёба(palatum durum) является вторичное костное твёрдое нёбо - нёбные отростки резцовых, верхнечелюстных костей и горизонтальные пластинки нёбных костей, срастающиеся по средней линии. Кости плотно покрыты слизистой оболочкой. За верхними резцами на твёрдом нёбе у большинства млекопитающих имеются парныеносо-нёбные отверстия, ведущие ввомеро-назальный (Якобсонов) орган(отсутствует у человека). По средней линии на нёбе проходит продольный шов, по сторонам от него – ряды поперечных валиков. Валики загнуты по направлению к глотке, их края могут сильно ороговевать. Максимально сильно такое ороговение выражено у усатых китов, где роговой слой эпителия нёбных валиков формирует цедильный аппарат - так называемый китовый ус. У животных, способных к отрыгиванию пищи, на задней части твёрдого нёба валики сглажены.

Мягкое нёбо(palatummolle) или нёбная занавеска является складкой слизистой оболочки, свисающей с заднего края твёрдого нёба. Внутри мягкого нёба имеются мышцы, сокращаясь, они укорачивают и поднимают нёбную занавеску, так, что при глотании она закрывает собой отверстия хоан. Край нёбной занавески –нёбная дужка– ограничивает сверху отверстие выхода из ротовой полости в глотку –зев. Форма, длина и степень натяжения нёбной занавески сильно различаются у разных млекопитающих. Так, если у крупного рогатого скота и свиней нёбная занавеска короткая (у последних снабжена «язычком») и в свободном состоянии не касается корня языка, то у лошадей, напротив, нёбная занавеска длинная, туго натянутая и плотно прилежит к корню языка, освобождая зев только при глотании. В толще мягкого нёба залегаетнепарная нёбная миндалина(см.), а также многочисленныенёбные слюнные железы.
Язык(lingua,s.glossa) млекопитающих – мощный мышечный орган, обладающий разнообразной подвижностью и очень высокой чувствительностью. Различаюткорень,телоиверхушку языка. Корень и тело языка имеют дорсальную и латеральные поверхности, вентрально прикрепляясь к мягким тканям дна ротовой полости. Верхушка языка имеет и вентральную поверхность, слизистая оболочка которой переходит на дно ротовой полости, образуяуздечку языка.
В толще языка, кроме мышц, залегает большое количество нервных окончаний, многочисленныепристенные слюнные железы(особенно в корне языка) инепарная язычная миндалина, расположенная почти на уровне зева. Кроме того, в корень языка входит язычный отросток подъязычной кости, представители некоторых видов млекопитающих имеют в языке хорошо развитый язычный хрящ (собаки, свиньи). Язык покрыт слизистой оболочкой с многослойным плоским эпителием, который на дорсальной поверхности языка может в разной степени ороговевать. На дорсальной и боковых поверхностях языка располагаются сосочки:механические –нитевидныеиконусовидные, они ороговевают и служат для продвижения пищи, ивкусовые сосочки для определения вкуса. У млекопитающих встречаются три вида вкусовых сосочков:грибовидные,валиковидныеилистовидные. Грибовидные сосочки представляют собой округлые слегка выпуклые образования, лежащие поодиночке на верхушке языка и группами на боковых поверхностях. Они несут на себе по несколько вкусовых луковиц. Валиковидные и листовидные сосочки, более крупные, располагаются соответственно на дорсальной и боковых поверхностях корня языка. Валиковидные сосочки имеют округлую или овальную форму, их возвышенная центральная часть окружена желобком. Листовидный сосочек представляет собой овальную пластинку, рассечённую несколькими поперечными щелями. В дно желобков (щелей) вкусовых сосочков впадают протоки слюнных желёз, а в стенках залегают многочисленные вкусовые луковицы, реагирующие на химический состав корма.
Дно ротовой полостиобразовано мягкими тканями. В его толще лежат мышцы, а также застенные и пристенные слюнные железы. К большей части дна ротовой полости прикреплён язык. По бокам от корня языка находятся складки, в которые открываются многочисленные протоки слюнных желёз. По бокам от уздечки языка у большинства млекопитающих имеются небольшие парные выросты слизистой оболочки –голодные бородавки, куда открываются протоки подчелюстной и подъязычной слюнных желёз.
Зубы(dentes) млекопитающих помещаются в альвеолах челюстных костей (текодонтные), одно и двусменные (моно- и дифиодонтные (см. выше)). Особенности зубной системы млекопитающих связаны с тем, что млекопитающие являются единственными позвоночными, способными длительное время обрабатывать пищу во рту – жевать. Такая способность появилась благодаря образованию вторичного твёрдого нёба: хоаны оказались сдвинуты каудально и ротовая полость потеряла связь с носовой (напомним, что амфибии, рептилии и птицы не могут дышать, когда ротовая полость занята пищей).
Зубная система млекопитающих гетеродонтная, то есть на зубной аркаде располагаются разные по форме, функции и сменности зубы. Передние зубы – резцы(dentes incisivi), за ними расположены по одному на челюстную костьклыки(dentes canini), далее –коренные зубы:премоляры(предкоренные, dentes premolares) имоляры(dentesmolares). У большинства млекопитающих резцы, клыки и премоляры являются дифиодонтными (двусменными, то есть первично вырастающие молочные (dentes decidui,Dd) заменяются на постоянные (dentes permanentes,Dp) зубы). Моляры млекопитающих – монофиодонтные. Молочные зубы млекопитающих вырастают тогда, когда челюсти имеют ещё очень маленькие размеры, а животное уже питается самостоятельно. Затем молочные зубы заменяются более крупными постоянными зубами. Моляры принадлежат молочной генерации, но по величине и форме соответствуют второй генерации премоляров.
Количество зубов разных видов и их распределение отображается зубной формулой. Существует два способа записи зубной формулы. Например, у взрослой свиньи она имеет вид: I 3/3,C1/1,P4/4,M3/3. Цифра в числителе показывает количество зубов на верхней челюсти (точнее, на её половине – на одной челюстной кости), в знаменателе – на нижней челюсти. При другом способе записи формула записывается как одна дробь, без упоминания видов зубов. При отсутствии каких-либо зубов пишется ноль.
П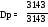 ример:
зубная формула свиньи:
ример:
зубная формула свиньи:
Зубная формула современных млекопитающих редуцирована (уменьшена). Наиболее полный вариант зубной формулы можно видеть у некоторых сумчатых, где в числителе будет стоять 5155, а в знаменателе - 4155. Причём все зубы, кроме первого премоляра, являются молочными. У ластоногих, грызунов и рукокрылых, напротив, угнетена молочная генерация, к ней принадлежат только моляры. Землеройки и некоторые грызуны совершенно лишены молочной генерации зубов. У зубатых китов количество зубов больше и они все внешне одинаковы, то есть система выглядит гомодонтной. Это результат вторичного упрощения зубной системы, связанного с образом жизни и питания. Считается, что множество простых конических зубов дельфина появились вследствие разделения многокорневых зубов. У муравьедов не формируется даже эмбриональная закладка зубов (как у птиц).
Кроме вертикальной смены зубов наблюдается и горизонтальная смена. Так, у слонов закладываются по 6 премоляров (3 молочных и 3 постоянных), но действует всегда только 1 зуб на каждой челюстной кости. Когда зуб стирается и выпадает, на его место сдвигается зуб, ранее располагавшийся каудально в десне. При этом перегородка между альвеолами разрушается. Такая же горизонтальная смена присуща ламантинам.
Коренные зубы млекопитающих различаются по форме жевательной поверхности, которая связана с типом питания животного. Так хищникам свойственны секодонтные(режущие)зубы, всеядным животным –бунодонтные(бугорчатые), травоядным -лофодонтныеиселенодонтные(складчатые и лунчатые). Исходный для млекопитающих тип поверхности зубов –трибосфеничекий(труще-режущий) – присущ насекомоядным, рукокрылым. Также с типом потребляемого корма связано формирование в классе млекопитающихкороткокоронковых(брахиодонтных) идлиннокоронковых(гипсодонтных) зубов. Короткокоронковый зуб имеет коронку, шейку и корень, при этом вся коронка находится над десной. Такие зубы бывают секодонтными и бунодонтными, они свойственны животным, потребляющим относительно мягкую пищу без длительного перетирания. По мере стирания пульпарная полость такого зуба заполняется дентином, а при полном стирании коронки короткокоронковый зуб обычно выпадает. Длиннокоронковый зуб состоит из длинной коронки и короткого корня. Он закладывается в челюстных костях сразу почти в полном размере (не сразу развивается корень зуба), а затем, в течение всей жизни, постепенно «выдвигается» на поверхность, так, что при правильном стирании над десной всегда видна часть коронки одной и той же высоты. Скорость выдвигания зуба определяется давлением: в норме при сомкнутых челюстях коренные зубы соприкасаются, а если они «отрастают» слишком длинными, давление усиливается и «рост» прекращается. Такие зубы свойственны животным, тщательно перетирающим грубую растительную пищу. Молочные зубы всегда являются короткокоронковыми. Отдельного внимания заслуживают так называемые «зубы с открытым корнем». Такие зубы действительно растут всю жизнь: на протяжении всей жизни животного сохраняются эмалевые клетки, продуцирующие поверхностный слой и рост зуба практически неограничен. Корень у постоянно растущего зуба не формируется вовсе. К таким зубам, например, относятся резцы грызунов и зайцеобразных, бивни (резцы) слонов и нарвалов (бивень нарвала - левый верхний резец), клыки свиней и др.
Дёсны(gingivae) – это слизистая оболочка, плотно покрывающая зубные края челюстных костей. Дёсны тесно обхватывают зубы и сливаются с периодонтом и надкостницей, они содержат много кровеносных сосудов и относительно мало нервных окончаний.
Слюнные железы(glandulae salivales) у млекопитающих чрезвычайно развиты в связи с длительной обработкой пищи в ротовой полости. Во всех мягких частях ротовой полости – в губах, щеках, языке, дне ротовой полости, в мягком нёбе – имеютсяпристенные слюнные железы, открывающиеся многочисленными протоками. В корне языка они составляют до ½ его массы. Кроме того, есть крупныезастенные железы, наиболее обычны из которых околоушная, подчелюстная и подъязычная.Околоушная слюнная железа(glandula parotis) лежит ниже наружного слухового прохода на каудолатеральном крае большой жевательной мышцы. Её проток на уровне лицевой сосудистой вырезки переходит с медиальной поверхности нижней челюсти на латеральную и открывается в преддверие ротовой полости на уровне верхних коренных зубов.Подчелюстная слюнная железа(нижнечелюстная железа, glandula mandibularis) располагается каудально в межчелюстном пространстве, иногда доходя до ярёмных отростков. Её проток открывается на дне ротовой полости рядом с уздечкой языка, в так называемых голодных бородавках.Подъязычная слюнная железа(glandula sublingualis) лежит в межчелюстном пространстве оромедиальнее подчелюстной железы. Проток одной её части сливается в конечном участке с протоком подчелюстной железы, другая часть имеет множественные протоки, открывающиеся по бокам от тела языка в дно ротовой полости.
Слюнные железы млекопитающих могут выделять как слизистый, так и серозный секрет (реже – клейкий секрет). В секрете серозных желёз имеются амилолитические ферменты (у жвачных отсутствуют).
Глотка(farinx) млекопитающих является относительно обособленной камерой. В ней происходит перекрёст пищеварительных и дыхательных путей. Входами в глотку являются: сверху – парныехоаны(из носовой полости), снизу –зев(из ротовой полости). Выходы: наверх – впищевод, вниз – вгортаньи далее в трахею. Таким образом, хотя млекопитающие и могут сколь угодно длительно обрабатывать пищу в ротовой полости и при этом ничто не мешает процессу дыхания, но во время глотания дыхательные пути должны быть перекрыты. Во время прохождения пищевого кома через глотку хоаны закрываются напрягающимся мягким нёбом, а отверстие гортани – одним из хрящей – надгортанником. (В глотательном движении важную роль играет подъязычный аппарат, сопрягающий движения корня языка и гортани.) Вне момента глотания мягкое нёбо расслаблено и свисает вниз по направлению к корню языка, хоаны открыты, как и гортань, а не имеющий твёрдой стенки пищевод спадается.
Верхняя часть глотки, по которой проходит воздух, выстлана однослойным многорядным мерцательным («дыхательным») эпителием и именуется носоглоткой. Нижняя часть глотки, относящаяся к пищеварительным путям, выстлана многослойным эпителием и называетсяротоглоткой. В носоглотке недалеко от хоан располагаются парные отверстияглоточно-барабанных (Евстахиевых) труб, соединяющих глотку с полостью среднего уха и служащих для выравнивания давления с внутренней стороны от барабанной перепонки.
Через глотку проходят и воздух и пища, она является местом весьма тесного контакта внутренней среды организма с внешней средой. Функцию защиты выполняют располагающиеся здесь многочисленные миндалины(tonzillae) – скопления лимфатических фолликулов. Миндалины выполняют двойную функцию: здесь происходит размножение лимфоцитов для защиты на местном уровне, а также «знакомство» молодых лимфоцитов с различными антигенами – обучение их для дальнейшей работы на организменном уровне. Миндалины образуют в глотке так называемоеглоточное лимфоэпителиальное кольцо, в его составе: непарные язычная, нёбная и глоточная, парные нёбные, околонадгортанные и трубные (у животных разных видов какие-то миндалины могут отсутствовать). На анатомических препаратах обычно хорошо видны парные нёбные миндалины, располагающиеся в местах прикрепления нёбной дужки к корню языка.
ПЕРЕДНЯЯ КИШКА
Пищевод(oesophagus) начинается от ротоглотки и служит для проведения пищи в нижние части пищеварительного аппарата, где происходит основная часть обработки их пищеварительными ферментами. Пищевод может иметь различную длину, в зависимости от наличия и расположения желудка, тем не менее, присутствует он всегда. Наличие пищевода позволяет изолировать среду желудка (или кишечника – при отсутствии желудка) от среды ротоглотки. При отсутствии желудка пищеводом именуется часть кишки до печёночного (желчного) протока.
Пищевод выстлан многослойным плоским эпителием, часто ороговевающим. У миног, взрослых рыб, амфибий и рептилий он содержит ресничные и бокаловидные клетки. Слизистая оболочка пищевода не содержит желёз, они могут располагаться в подслизистой основе. Секрет желёз пищевода не содержит пищеварительных ферментов за исключением случаев, когда железы конца пищевода вырабатывают пепсиноген (желудочный фермент).
В составе пищевода может формироваться слепой одинарный или двойной мешок – зоб. В нём происходит хранение и мацерация корма.
У рыб пищевод (редко – желудок) связан с плавательным пузырём, по крайней мере, на ранних этапах онтогенеза.
Желудок (gaster,s.ventriculus) располагается за пищеводом, может состоять из нескольких более или менее резко разделённых камер. Части многокамерного желудка могут значительно различаться морфологически и функционально. Железы желудка (или, по крайней мере, его основной части) у всех позвоночных вырабатывают желудочный сок сходного состава: в нём присутствуют соляная кислота и фермент пепсин, обеспечивающий распад крупных белковых молекул на олигопептиды. Вероятно, изначально желудок сформировался как камера для хранения и мацерации пищи у хищников.
Желудок, как правило, имеет хорошо развитую мышечную оболочку, часто с увеличенным количеством слоёв. При переходе в кишку мышечная оболочка формирует сфинктер (пилорический сфинктер).
Наиболее часто встречается у позвоночных серповидная форма желудка. Часть желудка при впадении пищевода называется кардиальной, у выхода кишки – пилорической, а средняя часть – фундальной (дно желудка). Вогнутая поверхность называется малой кривизной, выпуклая – большой.
Принцип строения трубкообразного органа
Все внутренние органы, имеющие форму трубки (или формирующиеся из органов, имевших форму трубки), имеют принципиально общую схему гистологического строения. В составе их стенок различают четыре оболочки.
Слизистая оболочка является самой внутренней, контактирующей с просветом органа. Она выстлана эпителием. Под эпителием располагается собственная пластинка слизистой оболочки из рыхлой соединительной ткани. Здесь проходят кровеносные сосуды и нервы, могут располагаться железы. На внешней границе слизистой оболочки часто располагается тонкий слой гладкомышечных клеток – мышечная пластинка слизистой оболочки.
Подслизистая основа образована рыхлой соединительной тканью, в ней находятся более крупные сосуды, чем в слизистой и могут располагаться концевые отделы желёз. При отсутствии мышечной пластинки слизистой оболочки граница между слизистой и подслизистой незаметна, в этом случае часто говорят об отсутствии подслизистой основы.
Мышечная оболочка преимущественно образована гладкой мышечной тканью, хотя иногда в ней наблюдается и поперечнополосатая (например, в составе пищевода). Мышечные клетки (или волокна) лежат двумя слоями: внутренний – с кольцевой ориентацией клеток, наружный – с продольной. Точнее, в обоих слоях клетки лежат по спирали, но с разной плотностью витков (соответственно, с разным углом наклона витков к оси). В различных органах и их участках количество слоёв мышечных клеток может быть от одного до трёх. Кольцевой слой мышечных клеток может формировать сфинктеры.
Наружный слой трубкообразного органа зависит от его топографии. У органов, не лежащих в полостях тела, наружный слой – адвентиция, рыхлая соединительная ткань, соединяющая орган с окружающими органами и тканями. Когда орган располагается в полости тела, он покрыт серозной оболочкой, состоящей из двух слоёв: собственной пластинки серозной оболочки (рыхлая соединительная ткань) и мезотелия (однослойный плоский эпителий мезодермального происхождения, висцеральный листок спланхнотома). Мезотелий обеспечивает свободное расположение органа в полости тела, не позволяя ему срастаться поверхностями с окружающими органами и тканями.
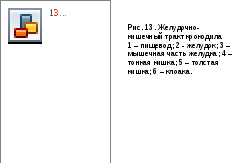
У ланцетникакишка, начинающаяся от резкого сужения в конце длинной глотки, короткая и практически не делится на отделы. Печёночный вырост находится сразу за глоткой – пищевода и желудка нет.
Пищеводкруглоротыхдлинный (в то время как глотка отсутствует), начинается от конца ротовой полости, идёт над дыхательной трубкой и продолжается до впадения желчного протока, где без внешних изменений переходит в кишку. Вдоль пищевода проходят высокие продольные складки. Эпителий пищевода содержит ресничные и бокаловидные клетки.
Желудоку круглоротых отсутствует.
У рыбпищеводкороткий, имеет продольные складки и сильно растяжим. Более длинный пищевод у рыб, не имеющих желудка (химеры, карповые и др.).
В начальной части пищевод костных рыб имеет связь с плавательным пузырём– либо проток, либо связка. В эпителии иногда имеются ресничные клетки (преимущественно у хрящевых рыб) и всегда естьодноклеточные слизистые железы. Как правило, в секрете желёз пищевода ферменты отсутствуют, хотя у рыб отдельных видов, заглатывающих целиком крупную добычу, они вырабатывают пепсиноген: тогда пища, не уместившаяся в желудке, начинает перевариваться в пищеводе.
Мускулатура пищевода рыб, как правило, поперечнополосатая.
Желудокрыб имеет три основных варианта формы: расширенная трубка, V-образный и V-образный с мешком в фундальной части. Железы желудка вырабатывают желудочный сок, только у двоякодышащих рыб железы в желудке отсутствуют. В пилорической части желудка имеется сфинктер, кардиального сфинктера у рыб нет, внешне место перехода пищевода в желудок слабо различимо. Складки желудка, в отличие от складок пищевода, извилистые или расположены косо. Мышечная оболочка желудка рыб развита хорошо, но, при этом, он весьма растяжим.

У амфибийпищеводболее длинный, чем у рыб, так как глотка значительно короче, а желудок сдвинут каудально лёгкими. В эпителии естьслизистые клетки, а у представителей некоторых видов эпителий реснитчатый. Пищевод имеет продольные складки и может сильно растягиваться.
Желудокамфибий серповидной формы, относительно слабо изогнут, имеет пилорический сфинктер. Железы желудка вырабатывают желудочный сок.

Длина пищеводарептилийсвязана с длиной шеи и размером лёгких, как правило, он имеет продольные складки и весьма растяжим. В эпителии имеются пристенные слизистые железы. У яичных змей, относящихся к ужовым, в пищевод дорсально вдаются сильно развитые вентральные гребни шейных позвонков, позволяющие разрушать скорлупу проглатываемых целиком яиц.
У всех рептилий желудокпредставляет собой вместительный мешковидный орган. Форма его связана с формой тела рептилии – от веретеновидной у змей до округлой у черепах. У крокодилов желудок имеет разделение на железистую и мышечную части (судя по обнаружению гастролитов у ископаемых останков динозавров и по современным родственникам крокодилов – птицам, это разделение, возможно, было свойственно всем архозаврам). В стенках железистого желудка залегают железы, вырабатывающие желудочный сок обычного состава. В мышечном желудке желёз нет, сокращение его мощных стенок и содержащиеся в нём камешки – гастролиты (имеющие экзогенное происхождение – животное их просто подбирает и заглатывает) обеспечивают измельчение корма, предварительно обработанного желудочным соком.
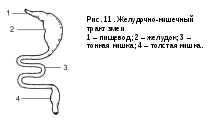





Пищеводптицвсегда очень длинный, а у видов, потребляющих пищу нерегулярно и в больших количествах (хищники, зерноядные), может иметь растяжимое образование –зоб, для временного запасания пищи перед ее перевариванием (а у зерноядных – и для мацерации). Зоб присутствует у соколообразных, курообразных, голубеобразных, попугаеобразных и некоторых ржанкообразных, деля пищевод на предзобную и зазобную части. У голубей, кроме того, эпителий зоба и пристенные зобные железы образуют секрет для выкармливания птенцов (так называемое «молочко»).
Желудокптиц разделен нажелезистую (proventriculus) имышечную(ventriculus)части, причем их относительное и абсолютное развитие различно. Железистая часть желудка предшествует мышечной, таким образом, пища, обработанная желудочным соком, затем подвергается механической обработке толстыми стенками и гастролитами в мышечной части. Какая-то одна часть желудка, или обе, велики, тонкостенны и растяжимы у птиц, глотающих целиком крупную добычу и лишенных зоба (бакланы, цапли, совы, чайки). У страуса часть железистого желудка лишена желез и выполняет функцию запасания корма, заменяя отсутствующий зоб. Мышечная часть желудка птиц выстлан изнутри кератиноподобнойкутикулой, выделяемой железами его эпителия. И кутикула, и мышечная стенка мышечной части желудка очень мощны у птиц, потребляющих грубую растительную пищу и, напротив, слабы у плотоядных. У последних в мышечной части желудка формируется погадка – отрыгиваемый затем комок из непереваренных остатков пищи. Погадки отрыгивают также и насекомоядные птицы.
Желудок у птиц однотипно расположен вблизи центра тяжести тела, что выгодно для животных, быстро меняющих положение тела в пространстве.



Пищеводмлекопитающихможет иметь различную длину в соответствии с различной длиной шеи. Дополнительно пищевод удлиняется за счёт развития грудо-брюшной преграды – диафрагмы, которая отодвигает желудок каудально. Эпителий пищевода многослойный плоский, иногда ороговевающий, в подслизистой основе имеются слизистые железы. Мышечная оболочка пищевода частично или полностью сформирована поперечнополосатой мышечной тканью, что особенно свойственно животным, способным к произвольному отрыгиванию.
Желудокмлекопитающих разных видов может иметь значительные различия в строении. Простейшая классификация делит желудки млекопитающих на однокамерные и многокамерные.
Однокамерный желудокмлекопитающего лежит почти поперёк тела животного, так, что кардиальная часть располагается слева, а пилорическая - справа. В целом желудок имеет форму полумесяца с более или менее расширенным и вытянутым телом желудка. Малая кривизна направлена краниодорсально, большая - каудовентрально. В кардиальной и пилорической частях желудка могут формироваться слепые мешки различного объёма. Пищевод обычно входит в желудок слегка расширяясь (свиньи) или без расширения (хищники). У эквидов (лошади, зебры) кольцевой и косой слои мускулатуры желудка формируют у пищеводного отверстия две петли, в сумме дающие мощныйсфинктер. При наполнении желудка пищей или газами сфинктер сжимается, так что рвотные движения для лошадей невозможны.
В большинстве случаев часть желудка (от 5 до 95 % площади поверхности начиная от отверстия пищевода) выстлана слизистой оболочкой пищеводного типа(имеет многослойный эпителий и не содержит желёз). У однопроходных желудок вовсе не имеет желёз, а у муравьедов их ничтожно мало (у ехидны и муравьеда низкийpHобеспечивается за счёт кислотности корма). Остальная площадь поверхности желудка выстланаслизистой оболочкой кишечного типа, с однослойным эпителием и железами, вырабатывающими желудочный сок. Весь цилиндрический эпителий желудка и часть желёз (кардиальные и пилорические) вырабатывают слизь, предохраняющую стенки желудка от самопереваривания. Мышечная оболочка желудка может содержать не два, а три, а местами даже четыре слоя (добавляются внутренний и наружный косые слои), что обеспечивает возможность сложных движений органа для перемешивания и сортировки содержимого.
К моногастричным(имеющим однокамерный желудок) относится большинство млекопитающих: хищники, грызуны, приматы, лошади и т.д. Типичные простые однокамерные желудки имеют человек, свинья, лошадь, собака, кошка.
Перед травоядными животными, потребляющими корма с высоким содержанием клетчатки, стоит серьёзная проблема, поскольку в организме позвоночных совершенно не вырабатываются целлюлозолитические ферменты. Такие ферменты, однако, вырабатываются некоторыми одноклеточными - инфузориями, бактериями. Поэтому травоядные (в меньшей степени – всеядные) млекопитающие имеют своего рода «мини-цеха» по переработки целлюлозы, где «работают» одноклеточные симбионты. Они играют и дополнительную роль, поставляя хозяину белок и витамины. Участки ЖКТ, где происходит микробиальное разложение целлюлозы, должны иметь большой объём и малоагрессивную среду. Эти условия могут быть выполнены либо в преджелудке, выстланном слизистой пищеводного типа, либо в толстом кишечнике. Таким образом, травоядные млекопитающие имеют либо сложный (многокамерный) желудок, либо объёмный толстый отдел кишечника (см. далее).
Особенность многокамерных желудковмлекопитающих (и коренное отличие от желудка птиц) заключается в том, что передние камеры желудка (преджелудок) служат для обработки пищи с помощью микроорганизмов (в первую очередь, инфузорий). Соответственно, преджелудок не содержит собственных (фундальных) желёз, вырабатывающих желудочный сок. Какая-то часть слизистой преджелудка может содержать кардиальные железы, вырабатывающие слизь. Только конечный отдел многокамерного желудка является истинным желудком, с собственными (фундальными) железами в слизистой оболочке и обычным составом желудочного сока.
К млекопитающим с многокамерным желудком – полигастричным- относятся, в первую очередь, жвачные парнокопытные. У них (овцы, козы, коровы, олени, антилопы) наблюдается максимальное количество камер желудка: 3 камеры преджелудка и истинный желудок. Сходные желудки у верблюдов и бегемотов.
Строение многокамерного желудка коровы.
Пищеводкоровы перед впадением в желудок расширяется, образуя воронку. Первая камера преджелудка –рубец(rumen) – самая большая. Её объём может составлять более сотни литров. Рубец разделён складками надорсальныйивентральный мешки, от которых каудально отходяткаудодорсальныйикаудовентральный слепые мешки. Эпителий рубца многослойный плоский, частично ороговевающий. Слизистая оболочка образуетсосочкидлиной до 1 см. В рубце имеется обильная микрофлора, представленная, в первую очередь, инфузориями. Здесь происходит расщепление клетчатки до летучих жирных кислот. К тому же, микрофлора рубца является дополнительным источником белка и витаминов.
Пищевод входит в дорсокраниальную часть рубца, называемую преддверием рубца. Оно сообщается широким отверстием со второй камерой –сеткой(reticulum). Слизистая оболочка сетки имеет невысокие складки, образующие сетчатый узор. Функция этой камеры, выстланнй слизистой пищеводного типа, - это разделение пищевых масс на фракции. Из сетки, через преддверие рубца крупная фракция ферментированного корма отрыгивается в ротовую полость, где пережёвывается. По дорсальной стенки сетки от отверстия пищевода проходитпищеводный (желудочный) желоб. Он служит для проведения молока, частично – воды, жидкого корма в истинный желудок (сычуг). Попадание молока в рубец привело бы к его створаживанию и образованию сгустка, переработать который рубец никак бы не смог. Пищеводный желоб вообще свойственен сложным желудкам, даже не относящимся к многокамерным (например, желудку кенгуру).
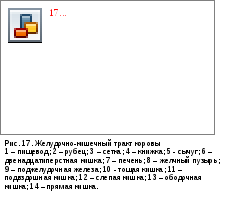
Третья камера преджелудка коровы называется книжка(omasum). Она сообщается с сеткой очень узким отверстием. Слизистая оболочка книжки формирует различной высоты складки –листочки. Листочки параллельны друг другу и лежат очень тесно. Эпителий образует на листочках ороговевающие сосочки. Книжка имеет толстую мышечную оболочку, пучки мышечных клеток от неё заходят в листочки. Функция книжки – перетирание и окончательное измельчение уже пережёванного корма. В ней корм, прошедший ферментацию в рубце и пережёванный, превращается в нежную пасту и переходит в сычуг.
Сычуг(abomasum) соответствует по строению однокамерному желудку. Он выстлан слизистой кишечного типа, с преобладанием фундальных желёз. В месте перехода сычуга в 12-перстную кишку имеется сфинктер.
Своеобразное строение имеют желудки кенгуру и некоторых обезьян (например, колобуса). Эти животные являются травоядными. Их желудки представляют собой «переходный вариант» от однокамерного к многокамерному желудку. Продольный слой мускулатуры этих желудков формирует тении- лентовидные утолщения, которые при нормальном тонусе собирают стенку очень удлинённого трубкообразного желудка в ряды глубоких карманов. Относительный объём желудка очень велик, большая часть его поверхности (начиная от кардиальной области) выстлана слизистой пищеводного типа. Наличие карманов позволяет хранить в кардиальной части желудка корм в относительной изоляции от желудочного сока, там возможны и процессы микробиальной обработки.
Желудки грызунов, хотя и не бывают истинно многокамерными, у многих видов разделены складками слизистой или имеют в кардиальной части относительно изолированные слепые мешки с пищеводной выстилкой. Это особенно характерно для животных, потребляющих большое количество грубого корма с низкой питательностью. Полёвки, хомяки и другие продолжают поиск и потребление легкоусвояемого, но редко встречающегося корма, пока грубый корм подвергается микробиальной обработке в слепом мешке.
Необычным примером многокамерного желудка является желудок дельфина – животного, потребляющего исключительно животную пищу. Пищевод входит в хорошо развитый преджелудок - слепой мешок со слизистой пищеводного типа. Рядом с отверстием пищевода находится вход в основную часть желудка. В свою очередь, она чётко подразделяется складкой слизистой на тело желудка с фундальными железами и пилорическую часть, железы которой вырабатывают только слизь. Считают, что здесь преджелудок выполняет функцию накопления пищи.
СРЕДНЯЯ И ЗАДНЯЯ КИШКА
Средняя кишка включает в себя тонкий отдел кишечника и связанные с ним застенные железы, задняя кишка – толстый отдел кишечника.
Передней границей тонкого отдела кишечникаслужит пилорический сфинктер желудка, при отсутствии желудка – место впадения желчного протока. Тонкий кишечник – место протекания большинства пищеварительных процессов, здесь осуществляется всасывание простых соединений в кровь и лимфу. В просвет кишечника изливается желчь (от печени) и панкреатический сок (от поджелудочной железы). Желчь создаёт щелочную среду и эмульгирует жиры, делая их доступными для воздействия ферментов – липаз. Панкреатический сок содержит огромное количество различных ферментов: трипсин, пептидазу, липазу, нуклеазу и другие – разлагающих на низкомолекулярные составляющие почти все органические вещества пищи. Таким образом обеспечивается полостное пищеварение в тонкой кишке. Эпителий кишечника цилиндрический, каёмчатый, то есть большая часть его клеток – энтероцитов - несёт на апикальных полюсах микроворсинки. Много и бокаловидных клеток, выделяющих слизь. В щёточной каёмке происходит пристеночное пищеварение, наиболее эффективное, по мнению многих исследователей. Большая площадь поверхности мембран микроворсинок создаёт оптимальные условия для работы большинства ферментов, а также для интенсивного всасывания продуктов переваривания. Результаты ферментного разложения органических веществ поступают в клетки эпителия, а затем – в кровеносные сосуды подлежащей соединительной ткани. Исключение составляют жиры: в просвете кишечника они распадаются до глицерина и жирных кислот, затем в клетках эпителия восстанавливаются до триглицеридов и формируют микроскопические капли (хиломикроны), поступающие в лимфатические капилляры. (Поэтому лимфа в сосудах, оттекающих от кишечника (хилус) непрозрачная и желтоватая, и сосуды эти называют млечными.)
Площадь внутренней поверхности тонкого кишечника у большинства позвоночных значительно увеличивается складками и кишечными ворсинками– выростами, в которых участвует эпителий и соединительная ткань собственной пластинки слизистой оболочки. У подножия ворсинок располагаютсякишечные крипты– простые трубчатые железы слизистой оболочки. Крипты выстланы тем же эпителием, что и ворсинки и выполняют, в целом, те же функции. В криптах происходит размножение клеток, затем они смещаются вверх по ворсинке и на её вершине гибнут и отпадают. Срок жизни клеток кишечного эпителия около двух-трёх суток.
Тонкий отдел кишечника может быть очень коротким (тогда часть его функций берёт на себя толстый отдел), а может превышать длину тела животного в десятки раз. За исключением самой передней части, весь остальной тонкий кишечник подвешен на длинной брыжейке и может формировать петли. Травоядные животные имеют относительно более длинный тонкий кишечник, чем плотоядные.
Печень(hepar) формируется и в онто- и в филогенезе по типу железы – как вентральный вырост стенки кишки. Её роль в процессе пищеварения – выделение желчи, эмульгирующей жиры. Но уже у ланцетника она принимает на себя другую, более важную и более сложную функцию – фильтрации крови (детоксикации, очистки, регуляции состава). В печень поступает кровь от органов пищеварения и селезёнки, несущая, кроме питательных веществ, и продукты распада, эндо- и экзогенные яды, микроорганизмы и т.д. Макрофаги печени улавливают и обезвреживают крупные частицы. Собственные клетки печени –гепатоциты(эпителий энтодермального происхождения) выполняют множество функций, таких как:
- переработка агрессивного аммиака в инертные мочевину и мочевую кислоту;
- переработка гемоглобина (из селезёнки, продукт распада эритроцитов) в желчные пигменты – билирубин и биливердин;
- депонирование глюкозы в виде гликогена;
- депонирование жира и т.д.
Желчь, выделяемая печенью, у большинства позвоночных скапливается в желчном пузыре, из которого выбрасывается при необходимости. Анатомические формы печени могут быть весьма разнообразны, даже в пределах одного класса. В большинстве случаев печень имеет несколько долей. У многих рыб печень просто занимает в полости тела всё доступное пространство. Размер печени у одного животного может значительно изменяться в связи с изменением питания.
Поджелудочная железа(glandula pancreas) возникает как скопление кишечных желёз для эффективного синтеза ферментов и их выведения в просвет кишки. Обычно представляет из себя компактный застенный орган, у низших позвоночных может быть рассеяна по стенке кишки или сливаться с печенью. Панкреатический проток открывается в кишку самостоятельно, рядом с желчным протоком, либо впадает в желчный проток. Кроме экзокринной имеется и эндокринная часть поджелудочной железы, она вырабатывает гормоны, регулирующие углеводный обмен.
Задняя кишка у позвоночных весьма разнообразна по форме, размерам и особенностям функции. У животных с малой длиной тонкого кишечника в толстом продолжаются процессы переваривания пищи ферментами поджелудочной железы в присутствии желчи, а также активное всасывание низкомолекулярных соединений. В основном, однако, в толстом отделе кишечника происходит всасывание воды и минеральных солей. У многих травоядных и всеядных позвоночных задняя кишка дополнительно увеличена и расширена – там происходят процессы расщепления клетчатки симбиотическими бактериями. Начиная с рептилий, на границе средней и задней кишки появляется слепой вырост стенки кишечника – слепая кишка. Она может быть и парной. Задняя кишка заканчивается анальным отверстием или впадает в копродеум клоаки. В конечном участке толстого кишечника млекопитающих – прямой кишке – происходит окончательное высушивание химуса и формирование каловых масс.
Стенка толстой кишки выстлана однослойным цилиндрическим эпителием с большим количеством бокаловидных клеток (одноклеточных слизистых желёз). Крипты развиты на всём протяжении кишки, а ворсинки отсутствуют. Характерным для толстого кишечника является наличие большого количества лимфоидных элементов, как в стенке, так и в брыжейке кишки.
У миксин,хрящевых рыб, некоторых костных рыб, амфибий, рептилий, птиц и однопроходных млекопитающих кишечник впадает вклоаку– камеру, общую для пищеварительной, выделительной и половой систем. Часть клоаки, собственно связанная с кишкой, называетсякопродеум. У остальных позвоночных кишка заканчиваетсяанальным отверстием.
У ланцетникаглотка после жаберных щелей резко сужается и переходит в кишечную трубку. Сразу за глоткой от вентральной стенки кишки отходит мешкообразныйпечёночный выступ. Кишка продолжается до анального отверстия, не дифференцируясь на отделы. Стенка кишки не образует каких-либо выступов или складок.
У круглоротыхжелудок отсутствует, поэтому сразу за пищеводом, лежащим в глоточной области (глотка разделена на пищевод и дыхательную трубку), начинается кишка.Печенькомпактна, лежит под кишкой сразу за сердцем, имеется желчный пузырь. Кишка не подразделена на отделы. На всём её протяжении стенка кишки формирует складку, в которой участвуют все оболочки, кроме серозной. Складка низкая, идёт продольно, слабо закручиваясь по спирали. Эта складка, вероятно, представляет собой прообраз спирального клапана рыб (см.далее). Слизистая оболочка кишки не образует ворсинок, в эпителии имеются реснитчатые клетки.
Открывается кишка анальным отверстием на поверхности тела (миноги), либо в клоаку (миксины).
У хрящевых рыби некоторыхкостно-хрящевых(осетровые) средняя кишка очень короткая. В неё впадают желчный проток от компактной крупнойпечении зачастую множественные тонкие панкреатические протоки.
Поджелудочная железаможет быть частично объединена с печенью, а частично – залегать в стенке кишки отдельными островками. Задний отдел кишечника представлена так называемойспиральной кишкой. При рассмотрении снаружи она выглядит как относительно короткая, толстая, прямая трубка. На вскрытой кишке видно, что её внутренняя поверхность формирует складку – спиральный клапан. Клапан одним краем спирально спускается по стенке кишки, а другим – свободным – обращён к центру просвета кишки. Таким образом, пища в спиральной кишке проходит не напрямую, а по длинному спиральному спуску. У многих видов складка спирального клапана настолько высокая, что образует «рулон» из широкой спирально закрученной ленты в просвете кишки и пища проходит в узких щелях между витками. Вопрос об отнесении спиральной кишки к среднему или заднему отделу остаётся открытым, но функционально она скорее соответствует средней кишке. Вероятно, спиральная кишка – это эксперимент природы по удлинению пути прохождения пищи в кишечнике (для более полного её использования). Однако, лучшим вариантом в эволюции пищеварительной системы оказалось простое удлинение кишечника. Хотя в состав спиральной складки кишки и входит мышечный слой, но управление движением химуса в обычно организованной кишке оказывается более эффективным.
От конечной части кишки у хрящевых рыб отходит пальцеобразный вырост – ректальная железа.Считается, что она принимает участие в солевом обмене.
Кишечная трубка хрящевых рыб впадает в клоаку.
У костистых рыбкишечник начинается от пилоруса желудка либо, при его отсутствии, от желчного протока.Печеньможет быть компактной, а может занимать всё доступное пространство полости тела.Поджелудочная железаобычно располагается над кишкой, впадая своим протоком несколько каудальнее печени, однако её части (а иногда и вся железа) могут оказываться в составе печени, желчного протока или стенки кишки. Многим рыбам (и не только костистым) свойственно наличие в самой передней части кишки – непосредственно за желудком –пилорических придатков. Они представляют собой либо пучки длинных червеобразных слепо замкнутых трубочек (треска), либо компактный шаровидный орган, на разрезе напоминающий губку. В настоящее время единственная доказанная функция пилорических придатков – увеличение площади внутренней поверхности кишки.
У костистых рыб средняя и задняя кишка морфологически неразличимы. Длина кишечника зависит от типа питания: у растительноядных рыб кишечник заметно длиннее, его длина может в несколько раз превосходить длину тела. Стенка кишки может формировать невысокие продольные и/или поперечные складки, края которых могут быть неровными – с зубцами и выемками. Такие неровные края складок считаются предшественниками ворсинок. Настоящих ворсинок на стенке кишки у рыб нет. Имеются крипты и, в начальном участке кишки, железы в подслизистой основе.
У костистых рыб кишка заканчивается анальным отверстием.
Кишечник взрослых амфибийкороткий, чётко подразделяется на тонкий и толстый отделы. (У головастиков, которые питаются растительной пищей, кишечник длиннее, чем у взрослых животных.)Тонкая кишкаобразует несколько петель, на дорсальной поверхности ей начального участка лежит недлинная лентовиднаяподжелудочная железа.Печеньамфибий компактная, имеет несколько долей.Толстая кишкапрямая, имеет малую длину (около 1/5 длины тела), а по толщине превышает тонкую кишку приблизительно в3 раза.
Кишечник всех рептилийчетко подразделен натонкий и толстый отделы.В тонком отделе преимущественно идут процессы переваривания и всасывания, туда открываются протокипеченииподжелудочной железы, а в толстом кишечнике происходит формирование каловых масс. Кроме того, у растительноядных черепах и ящериц в месте перехода тонкой кишки в толстую появляется относительно небольшой слепо замкнутый мешок, стенка которого может образовывать складки на внутренней поверхности. Таким образом, у рептилий впервые наблюдаетсяслепая кишка, в которой, по-видимому, происходит бактериальная обработка пищи. Относительная длина кишечника, разумеется, больше у растительноядных рептилий, и в пределах кишечника у них толстый отдел превосходит тонкий. Толстый кишечник открывается в копродеум клоаки
Кишечник птицподразделяется натонкийитолстый отделы. К тонкому относятсядвенадцатиперстная кишка, уложенная петлей почти до самого конца полости тела,тощая – самая длинная – иподвздошная кишки. В петле двенадцатиперстной кишки в виде трёх-четырёх узких долей лежит поджелудочная железа.Печень, проток которой открывается в двенадцатиперстную кишку, может иметьжелчный пузырь, а у некоторых видов он отсутствует (голуби, страусы). Расположение печени однотипно, и она имеет вдавления от сердца, железистого и мышечного отделов желудка и двенадцатиперстной кишки. К толстому отделу кишечника относятсяободочнаяи параслепых кишок(прямой кишке соответствует у птиц копродеум клоаки). Слепые кишки лежат вдоль подвздошной, верхушками направлены вперёд. В составе каждой слепой кишки различаются толстая часть (дистальная) и тонкая – начальная часть (проксимальная). В слепых кишках птиц, в отличие от млекопитающих, весьма интенсивно идут процессы переваривания и всасывания, что подтверждается наличием на их эпителии микроворсинок. У большинства плотоядных птиц слепые кишки отсутствуют (аистообразные, пеликанообразные, соколообразные), что затрудняет обнаружение границы между тонким и толстым кишечником, и даже у зерноядных (голуби) могут быть очень малы.
У основания слепых кишок у птиц располагается скопление лимфоидной ткани, так называемая кишечная миндалина. Каудально от основания слепых кишок лежит ободочная кишка. У большинства птиц она короткая и тоньше двенадцатиперстной (кроме страуса, у которого она служит для микробиальной обработки грубого растительного корма и является самой длинной и самой толстой кишкой). Слепые кишки выполняют ту же функцию и достаточно велики у растительноядных (но не обязательно зерноядных) птиц – гусей, страусов, тетеревиных.
Кишечник большинства млекопитающихдлинный (многократно превышает длину тела животного) и чётко разделён на тонкий и толстый отделы, особенно у всеядных и растительноядных видов. В составе тонкого отдела кишечника морфологически выделяются двенадцатиперстная, тощая и подвздошная кишки, в составе толстого – слепая, ободочная и прямая.
Двенадцатиперстная кишка(intestinum duodenum) имеет относительно небольшую длину, она подвешена на короткой брыжейке и её расположение у представителей разных видов принципиально сходно. От пилоруса желудка она идёт к печени и делает у её воротS-образный изгиб. Затем направляется каудально, у некоторых видов доходя до уровня подвздошных костей, поворачивает вперёд, проходит под правой почкой и идёт вентрально, где переходит в тощую кишку. В петле 12-перстной кишки располагаетсяподжелудочная железа, которая у млекопитающих может иметь до трёх частей: тело, правую и левую доли. Проток поджелудочной железы впадает в кишку в районе S-образного изгиба, недалеко от желчного протока.
В печенимлекопитающих различают несколько долей, сильнее или слабее разделённых вырезками. У большинства млекопитающих естьжелчный пузырь, исключением являются эквиды. Печень располагается в куполе диафрагмы, её выпуклая поверхность называетсядиафрагмальнаяили париетальная, а вогнутая –висцеральная. На висцеральной поверхности находятся ворота печени, в которые входят печёночная артерия, воротная вена, нервы, а выходитпечёночный проток. Соединяясь спузырным протоком, он становитсяжелчным. Печёночные вены впадают непосредственно в каудальную полую вену, там, где она оставляет вдавление на дорсальном (тупом) крае печени. На печени оставляют вдавления желудок, пищевод, правая почка.
Концом 12-перстной и началом тощей кишки считается место, где кишка поворачивает вниз, брыжейка удлиняется и кишка собирается в складки. Тощая кишка(intestinumjejunum) имеет большую длину и слабую наполненность. В конечном участке тонкого кишечника складки пропадают, кишка выпрямляется и называется здесь ё.
На всём протяжении тонкого кишечника его слизистая оболочка формирует ворсинки и крипты. В 12-перстной кишке ворсинки неровные, слизистая формирует складки, которые в дальнейшем пропадают и ворсинки образуют ровный слой. На протяжении 12-перстной кишки и начального участка тощей имеются сложные железы в подслизистой основе, в других частях кишечника такие железы отсутствуют. В брыжейке тощей кишки имеются многочисленные лимфатические узлы и их скопления.
Подвздошная кишка(intestinumileum) обычно впадает в толстый кишечник на границе слепой и ободочной кишок, но иногда она впадает непосредственно в слепую кишку. Тогда выход из слепой в ободочную кишку находится рядом. На входе подвздошной кишки в толстую имеется складка стенки кишки, называемаяилеоцекальный (подвздошно-слепокишечный) клапан.
Толстый отдел кишечника начинается слепой кишкой(intestinumcaecum). Она и следующая за нейободочная кишка(intestinumcolon) особенно хорошо развиты у растительноядных моногастричных млекопитающих. У последних в толстом кишечнике происходит бактериальное расщепление клетчатки. Увеличение объёма и площади внутренней поверхности обеспечивается за счёт наличия тений и кармашков. Тении – это лентовидные утолщения продольного слоя мышечной оболочки, которые собирают стенку кишки в складки – кармашки. Обычно имеются 2-3 тении.
Длина толстого кишечника меньше, чем тонкого, но всё же превышает длину тела, иногда до 4 раз. Объём же толстого кишечника растительноядных моногастричных млекопитающих может быть значительно больше, чем объём тонкого. Кишки толстого отдела, особенно ободочная кишка, бывают уложены сложными петлями или спирально. Примером здесь является толстый кишечник лошадей и зайцеобразных, где происходит бактериальное переваривание клетчатки. Зайцеобразные имеют и возможность использования бактериального белка, которой лишены лошади. Им свойственна копрофагия– поедание собственных экскрементов. Обычные экскременты содержат гораздо больше вредных для организма веществ, чем полезных. Но у кроликов выделяется два вида экскрементов – дневные, обычные, в виде шариков, и ночные – в виде зелёных колбасок. Эти последние производятся слепой кишкой и через ободочную проходят быстро, именно они и поедаются животными, служа прекрасным источником легкопереваримого бактериального белка и витаминов.
У всеядных млекопитающих (человек, крыса) толстый кишечник имеет среднее развитие, а у плотоядных (кошка, норка, дельфин) – в разной степени упрощён или даже редуцирован.
Прямая кишка (intestinum rectum), как явствует из её названия, идёт прямо под позвоночником из поясничной области в тазовую полость и до ануса. В прямой кишке происходит всасывание воды и минеральных солей и окончательное формирование каловых масс. Степень высушивания кала может быть очень различной у животных разных видов.


Толстый кишечник имеет типичное для кишечника строение стенки. Отличается от тонкого кишечника отсутствием ворсинок, эпителий крипт более чем на 50 % представлен бокаловидными клетками. Для кишок толстого отдела характерно наличие большого числа лимфатических фолликулов в стенке. Лимфоциты из фолликулов выходят на внутреннюю поверхность слизистой оболочки. Верхушка слепой кишки у некоторых видов (человек, зайцеобразные) формирует слепой отросток – аппендикс. Он имеет очень узкий просвет и в стенках его лимфатические фолликулы лежат так плотно, что аппендикс называют «кишечной миндалиной».
Конец прямой кишки выстлан многослойным плоским эпителием, в его стенках открываются протоки анальных желёз кожного происхождения. Заканчивается прямая кишка анальным сфинктером (у однопроходных – впадает в копродеум клоаки).