

obshaja_cytologija
.pdfзамкнутые циклы. У E. coli периметр такого кольца составляет около 1,6 мм, и считается, что на клетку приходится одна гигантская циклическая молекула ДНК, одна бактериальная хромосома или генофор с молекулярной массой 4 х 109 Д. Самая маленькая бактериальная хромосома обнаружена в клетках
-------
* Единицы измерения ДНК: пг – 1 х 10-12 г; Да – 1,67-24 г; 1 нуклеотидная пара (н.п.) – 1 х 103 Д, величина н.п. 0,34 нм.
микоплазмы – 0,25 мм (для сравнения, длина ДНК на одну хромосому эукариотической дрожжевой клетки составляет около 4,6 мм, а у человека – 40
мм(!) в 1 хромосоме).
Уряда бактерий, например у B. subtilis имеется от 2 до 9 одинаковых молекул ДНК и соответственно несколько нуклеоидов. В других случаях (Azotobacter vinelandii) около 40 хромосом организованы в один нуклеоид.
С помощью авторадиографической методики было также обнаружено, что репликация такой кольцевой хромосомы у E. coli начинается на одной исходной (origin) точке репликации, образуются две репликационные вилки, которые по мере синтеза ДНК движутся вдоль молекулы до терминальной, конечной точки (рис. 21). Тем самым вся такая гигантская молекула ДНК представляет единицу репликации, репликон. Скорость репликации у бактерий составляет около 30 мкм в мин., что согласуется со временем удвоения клеток равным около 40 мин.
Бактериальные хромосомы всегда связаны с плазматической мембраной через специфические мембранные белки, которые взаимодействуют с ДНК в зоне старта ее синтеза (рис. 22). В процессе клеточного деления существенных изменений в компактности нуклеоплазмы не наблюдается, в отличие от эукариотических хромосом.
Такие кольцевые молекулы ДНК бактерий были получены при полном удалении белков (депротеинизация). Если же изучать выделенные целые нуклеоиды бактерий, то они представляют собой тела, состоящие из многочисленных суперспирализованных петель ДНК, отходящих от плотной
71
центральной области (рис. 23). В одну такую петлю или домен входит до 10-15 мкм ДНК, или около 40 000 н.п., а всего таких петель – около 120.
Обработка выделенных нуклеоидов РНК-азой и протеолитическими ферментами приводит к разрыхлению центральной области нуклеоидов, а короткая обработка ДНК-азой – к снятию сверхспирализации петель, и декомпактизации всего нуклеоида. Таким образом было показано, что компактизация нуклеоида связана с наличием связок, содержащих РНК и некоторые белки. Тем самым гигантская кольцевая молекула – хромосома с помощью РНК и белков многократно складывается, образуя многочисленные петли, ДНК которых подвергается сверхспирализации, что приводит к значительной компактизации всего комплекса, который и представляет собой нуклеоид. Степень компактизации ДНК в нуклеоиде бактерий достигает 1000 крат, а концентрация ДНК доходит до 10 мг/мл (!). Необходимо подчеркнуть, что часть ДНК нуклеоида связана с небольшим числом специальных основных белков, отличных от гистонов эукариот. Одна молекула одного из таких белков (H-NS) приходится на 400 н.п. ДНК. С петлями ДНК нуклеоида связано большое число молекул различных синтезируемых РНК и рибосом, которые обнаруживаются по периферии нуклеоплазмы.
Одна из моделей организации нуклеоида предполагает, что центральная его часть представлена неактивной и сверхспирализованной ДНК, тогда как по его периферии расположены деспирализованные петли, на которых происходит синтез различных РНК (рис. 24).
Отличительной чертой ядерных структур прокариот является то, что у них синтез РНК и синтез белка может происходить одновременно: рибосомы связываются с еще не до конца синтезированными молекулами иРНК и производят на них синтез белка. Таким образом возникает тройственный синтетический комплекс: ДНК – синтезирующая цепь РНК – рибосомы с синтезируемой полипептидной цепочкой (рис. 25). Такая ситуация возможна лишь в том случае, когда образующаяся молекула иРНК не подвергается
72
дальнейшей модификации типа процессинга, характерного для эукариотических клеток (см.ниже). У прокариотов, таким образом, процессы транскрипции и трансляции не разобщены территориально, в то время как у эукариотических клеток эти процессы протекают в двух разных компартментах, разделенных специальной ядерной оболочкой.
Отличается поведение ядерного материала прокариотов от такового эукариот при делении клетки, и в течение клеточного цикла.
Клеточный цикл – это время существования клетки от деления до деления. Как уже указывалось, деление всех типов клеток происходит только после удвоения ДНК. У бактерий часто сам процесс разделения тела клетки, цитотомия, не связана с окончанием синтеза ДНК, т.к. до наступления клеточного деления может начаться второй или даже третий раунд репликации ДНК. В результате такого беспрерывного синтеза ДНК в быстро растущих культурах на каждую разделившуюся клетку приходится одна кольцевая хромосома на промежуточных стадиях ее дальнейшего удвоения (рис. 26), т.е. каждая дочерняя клетка сразу после деления уже содержит частично реплицированный геном.
При делении бактериальных клеток не происходит особой конденсации ДНК в составе нуклеоида. По мере роста клетки в длину зона нуклеоида после синтеза ДНК увеличивается, а затем делится с помощью специального механизма. Обособление и разъединение двух дочерних хромосом связано с расхождением мест прикрепления хромосом к плазматической мембране (см. ниже).
Ядро эукариотических клеток
Сам термин «ядро» впервые был применен Брауном в 1833 г. Для обозначения шаровидных постоянных структур в клетках растений. Позднее такую же структуру описали во всех клетках высших организмов.
Ядерный аппарат эукариотических клеток имеет ряд отличий от прокариотических. Во-первых, ДНК-содержащий компонент отделен от
73
цитоплазмы специальной оболочкой (ядерная оболочка), во-вторых, количество ДНК в ядах эукариот в тысячи раз больше, чем в составе нуклеоидов бактерий, в-третьих, ДНК эукариот представляет собой сложный нуклеопротеидный комплекс, образующий специальную структуру – хроматин, из которого и состоят эукариотические хромосомы. Далее – в состав ядер эукариот входят несколько физически не связанных хромосом, каждая из которых содержит одну линейную гигантскую молекулу ДНК. Каждая хромосомная ДНК представляет собой полирепликонную структуру, т.е. содержит множеств автономно реплицирующихся участков. Синтез и образование транскриптов эукариотических клеток сопровождается процессами вторичной их перестройки, «созревания», включающих в себя как фрагментацию (процессинг), так и сращивание отдельных фрагментов ДНК (сплайсинг). Наконец, в ядрах не происходит синтеза белков, т.е. в эукариотических клетках процессы синтеза ДНК и РНК разобщены от процесса синтеза белков.
Клеточное ядро, обычно одно на клетку (есть примеры многоядерных клеток), состоит из ядерной оболочки, отделяющей его от цитоплазмы, хроматина, ядрышка и других продуктов синтетической активности, ядерного белкового остова (матрикса) и кариоплазмы (или ядерного сока) (рис. 27.). Эти основные компоненты встречаются практически во всех неделящихся клетках эукариотических одноили многоклеточных организмов.
Главный компонент ядер, хроматин, является структурой, выполняющей генетическую функцию клетки, в хроматиновой ДНК заложена практически вся генетическая информация. Ядерная оболочка выполняет сложную барьернорецепторную, а также транспортную и каркасную функцию. Нехроматиновый ядерный белковый остов (матрикс) обеспечивает не только пространственное расположение хромосом в ядре, но и участвует в их функциональной активности. Одним из хромосомных участков, определяющих синтез рРНК и образование клеточных рибосом, является ядрышко. Кроме того в ядре в связи с хроматином и матриксом обнаруживаются различные рибонуклеопротеидные
74
структуры, содержащие разные типы РНК. Между всеми этими компонентами заключена жидкая фаза клеточного ядра, кариоплазма, в которой протекают многие процессы, связанные как с ядерным метаболизмом, так и с внутриядерным транспортом белков и РНК.
При наблюдении многих живых клеток, особенно растительных или же клеток после фиксации и окраски, внутри ядра выявляются зоны плотного вещества, которое хорошо воспринимает разные красители, особенно основные. Благодаря такой способности хорошо окрашиваться этот компонент ядра и получил название «хроматин» (Флемминг, 1880). Способность хроматина воспринимать основные (щелочные) красители указывает на его кислотные свойства, которые определяются тем, что в состав хроматина входит ДНК в комплексе с белками. Такими же свойствами окрашиваемости и содержанием ДНК обладают и хромосомы, которые можно наблюдать во время митотического деления клеток.
Вотличие от прокариотических клеток, ДНК-содержащий материал хроматина эукариот, может пребывать в двух альтернативных состояниях: деконденсированном в интерфазе и в максимально уплотненном – во время митоза, в составе митотических хромосом.
Внеделящихся (интерфазных) клетках хроматин, выявляемый в обычный микроскоп, может равномерно заполнять объем ядра или же располагаться отдельными сгустками (хромоцентры). Часто он особенно четко выявляется на периферии ядра (пристеночный, маргинальный, примембранный хроматин) или образует внутри ядра переплетения довольно толстых (около 0,3 мкм) и длинных тяжей, образующих подобие внутриядерной сети. Такие ядра часто встречаются в клетках растений (рис. 28, 29, 30).
Хроматин интерфазных ядер представляет собой несущие ДНК тельца (хромосомы), которые теряют в это время свою компактную форму, разрыхляются, деконденсируются. Степень такой деконденсации хромосом может быть различной в ядрах разных клеток. Когда хромосома или ее участок
75
полностью деконденсирован, тогда эти зоны называют диффузным хроматином. При неполном разрыхлении хромосом в интерфазном ядре видны участки конденсированного хроматина (иногда называемого гетерохроматин). Показано многочисленными работами, что степень деконденсации хромосомного материала, хроматина, в интерфазе может отражать функциональную нагрузку этой структуры. Чем более дифффузен хроматин интерфазного ядра, тем выше в нем синтетические процессы. Так, в клетках лимфоцитов хроматин образует значительные скопления по периферии клеточного ядра. При стимуляции этих клеток к синтезу ДНК по мере включения предшественника ДНК 3Н-тимидина происходит постепенная деконденсация хроматина. Таким же образом меняется структура хроматина при синтезе РНК. Падение синтеза ДНК и РНК в клетках обычно сопровождается увеличением зон конденсированного хроматина. Так, в эритроцитах низших позвоночных практически весь хроматин ядер находится в конденсированном состоянии, и в этих ядрах не происходит синтеза ни РНК, ни ДНК. Если же ядра этих клеток стимулировать к синтезу РНК, например, в гетерокарионах (см. ниже), то они переходят в диффузное состояние.
Максимально конденсирован хроматин во время митотического деления клеток, когда он обнаруживается в виде телец – хромосом. В этот период хромосомы не несут никаких синтетических нагрузок, в них не происходит включения предшественников ДНК и РНК.
Исходя из этого можно считать, что хромосомы клеток могут находиться в двух структурно-функциональных состояниях: в рабочем, частично или полностью деконденсированном, когда с их участием в интерфазном ядре происходят процессы транскрипции и редупликации, и в неактивном – в состоянии метаболического покоя при максимальной их конденсации, когда они выполняют функцию распределения и переноса генетического материала в дочерние клетки.
76
Эухроматин и гетерохроматин
Было отмечено многими исследователями, что степень структуризации, конденсации хроматина в интерфазных ядрах может быть выражена в разной мере. Так в интенсивно делящихся и в мало специализированных клетках ядра имеют диффузную структуру, в них кроме узкого периферического ободка конденсированного хроматина встречается небольшое число мелких хромоцентров, основная же часть ядра занята диффузным, деконденсированным хроматином. С другой стороны в клетках высокоспециализированных или в клетках, заканчивающих свой жизненный цикл, хроматин представлен в виде массивного периферического слоя и крупных хромоцентров, блоков конденсированного хроматина. Такую структуру имеют, например, ядра нормобластов (одна из стадий дифференцировки эритроцитов), ядра зрелых лейкоцитов. Эти два примера могут иллюстрировать общее правило: чем больше в ядре доля конденсированного хроматина, тем меньше метаболическая активность ядра. При естественной или экспериментальной инактивации ядер происходит прогрессивная конденсация хроматина, и, наоборот, при активации ядер увеличивается доля диффузного хроматина.
Однако при метаболической активации не всякие участки конденсированного хроматина могут переходить в диффузную форму. Еще в начале 30-х годов было замечено Э. Гейтцем, что в интерфазных ядрах существуют постоянные участки конденсированного хроматина, наличие которого не зависит от степени дифференцированнности ткани или от функциональной активности клеток. Такие участки получили название гетерохроматина, в отличие от остальной массы хроматина – эухроматина (собственно хроматина). По этим представлениям, гетерохроматин – компактные участки хромосом, которые в профазе появляются раньше других частей в составе митотических хромосом, и в телофазе не деконденсируются, переходя в интерфазное ядро в виде интенсивно красящихся плотных структур (хромоцентры). Первоначально понятие гетерохроматина имело сугубо морфологическое значение, потому что,
77
изучая препараты окрашенных ядер, конечно нельзя знать, может ли данный участок конденсированного хроматина, хромоцентр, перейти в будущем в разрыхленное, эухроматическое состояние, или нет. В связи с этим в специальной цитологической литературе часто без всякого основания любой участок конденсированного хроматина стали называть гетерохроматином. Процесс же общей конденсации хроматина, например в ядрах лейкоцитов, называли гетерохроматизацией ядер. На самом же деле в составе ядерного хроматина только лишь некоторые участки практически никогда не теряют особого конденсированного состояния. Такими постоянно конденсированными зонами чаще всего являются центромерные и теломерные участки хромосом. Кроме них постоянно конденсированными могут быть также некоторые участки, входящие в состав плечей хромосом – вставочный или интеркалярный гетерохроматин, который в ядрах также представлен в виде хромоцентров. Такие постоянно конденсированные участки хромосом в интерфазных ядрах сейчас принято называть конститутивным (постоянным) гетерохроматином. Здесь же необходимо отметить, что участки конститутивного гетерохроматина обладают целым рядом особенностей, которые отличают его от остального хроматина. Конститутивный гетерохроматин генетически не активен, он не транскрибируется, реплицируется он позже всего остального хроматина, в его состав входит особая (сателлитная) ДНК, обогащенная высокоповторяющимися последовательностями нуклеотидов (см. ниже); он локализован в центромерных, теломерных и интеркалярных зонах митотических хромосом. Доля конститутивного хроматина может быть неодинаковой у разных объектов. Так у млекопитающих на него приходится 10-15% всего генома, а у некоторых амфибий – даже до 60%. Функциональное значение конститутивного гетерохроматина до конца не выяснено, предполагается, что он несет ряд важных функций, связанных со спариванием гомологов в мейозе, со структуризацией интерфазного ядра, с некоторыми регуляторными функциями.
78
Вся остальная, основная масса хроматина ядра может менять степень своей компактизации в зависимости от функциональной активности, она относится к эухроматину. Эухроматические неактивные участки, которые находятся в конденсированном состоянии, стали называть факультативным гетерохроматином, подчеркивая необязательность такого его состояния. Хорошим примером факультативного гетерохроматина может служить X- хромосома в организме человека. В клетках мужской особи X-хромосома деконденсирована, она активна, транскрибируется и морфологически не выявляется из-за своего рыхлого, диффузного состояния. В клетках женского организма, где присутствуют две X-хромосомы, одна из них находится в активном, диффузном состоянии, а вторая – в неактивном, конденсированном, она временно гетерохроматизована. В этом состоянии она может существовать в течение всей жизни организма. Но потомки ее, попадая в клетки мужского организма следующего поколения, снова будут активированы.
Вдифференцированных клетках всего лишь около 10% генов находится в активном состоянии, остальные гены инактивированы и, соответственно, находятся в составе конденсированного хроматина (факультативный гетерохроматин). Это обстоятельство объясняет, почему большая часть хроматина ядра структурирована.
Втаблице 1. даны сравнительные общие характеристики эухроматических и гетерохроматических районов интерфазных хромосом.
Таблица 1.
79
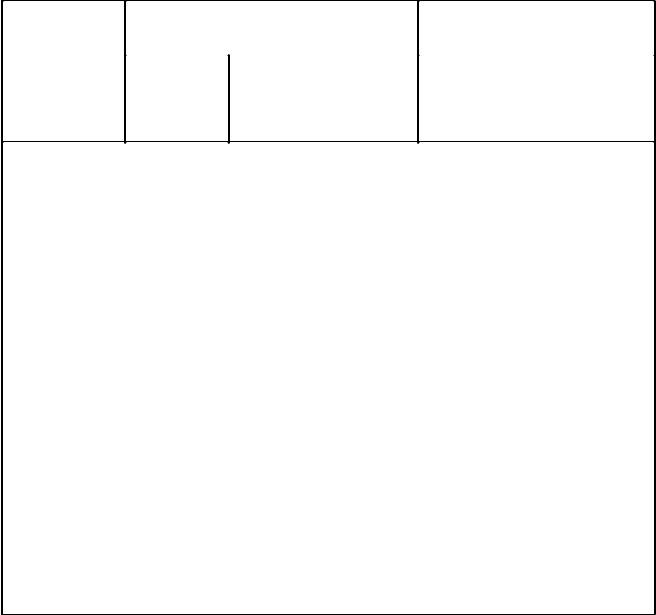
|
|
Эухроматин |
|
Гетерохроматин |
Свойства |
|
|
конститутивный |
|
|
|
Неактивный |
|
|
|
Активный (факультативный |
|
||
|
|
гетерохроматин) |
|
|
Структура |
диффузный |
конденсированный |
конденсированный |
|
Синтез РНК |
+ |
- |
- |
|
Синтез ДНК |
+ |
+ |
+ |
|
|
|
|
|
поздняя репликация |
Тип нуклеотид- |
уникальные, |
уникальные, |
высокоповторяющаяся, |
|
ных последова- |
умеренные |
умеренные |
сателлитная ДНК |
|
тельностей ДНК повторы |
повторы |
|
||
Локализация |
плечи |
плечи |
центромера, теломера, |
|
|
|
хромосом |
хромосом |
интеркалярный гетеро- |
|
|
|
|
хроматин |
Хромосомный цикл
Как известно, половые женские и мужские клетки несут одинарный набор хромосом и, следовательно, содержат в 2 раза меньше ДНК, чем все остальные клетки организма. Половые клетки (сперматозоиды и ооциты) с одинарным набором хромосом называют гаплоидными. Плоидность (от греч. ploos – кратность) обозначают буквой n, так, клетки с 1n – гаплоидны, с 2n – диплоидны, с 3n – триплоидны и т.д. Соответственно количество ДНК на клетку
(с) зависит от ее плоидности: клетки с 2n числом хромосом содержат 2с количества ДНК. При оплодотворении происходит слияние двух клеток, каждая
80