6. Репликация рнк. Репликация геномов ретровирусов. Обратная транскриптаза. Репликация рнк с образованием рнк.
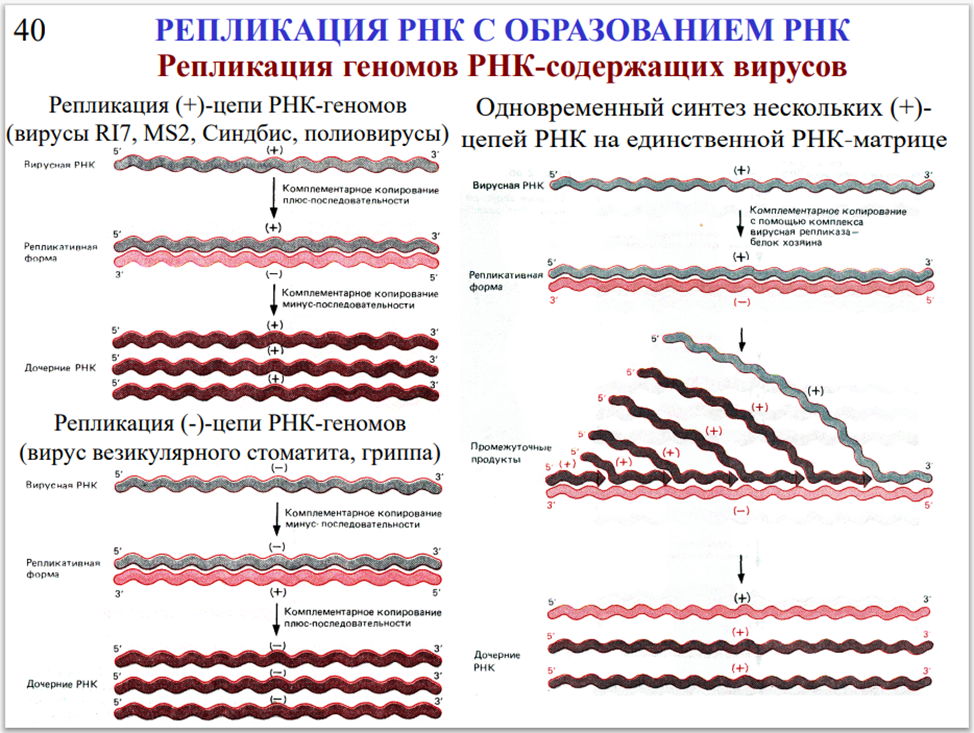
Процесс реализации генетической информации обычно носит однонаправленный характер. При транскрипции ДНК образуются молекулы РНК; исключение составляет обратная транскрипция у ретровирусов при их попадании в клетку. Полипептиды нельзя рассматривать в качестве носителей генетической информации; трансляция РНК в полипептид всегда носит необратимый характер. Описываемые механизмы обладают одинаковой эффективностью как для генетической информации прокариот и эукариот, так и для информации, содержащейся в вирусах. Геномы всех клеточных организмов представляют собой ДНК-дуплексы. Геномы вирусов могут состоять из ДНК или РНК, причем независимо от типа нуклеиновой кислоты геном вируса может быть как двух-, так и одноцепочечным (дц и оц). Детали механизма репликации нуклеиновых кислот варьируют в различных вирусных системах, однако все они базируются на принципе комплементарности.
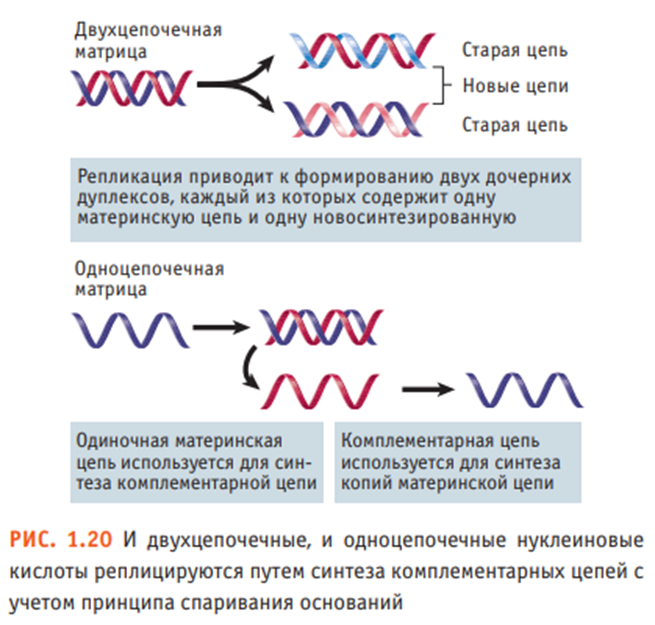
Клетки воспроизводят свою геномную ДНК по механизму полуконсервативной репликации. Двухцепочечные геномы РНК- и ДНК-содержащих вирусов также реплицируются с использованием отдельных цепей дуплекса в качестве матрицы для синтеза комплементарной цепи. Вирусы, несущие одноцепочечные геномы, для синтеза комплементарной цепи используют свою единственную цепь в качестве матрицы. А уже новая, комплементарная исходному геному цепь, в свою очередь, используется для синтеза молекул, идентичных оригинальной матрице. В процессе вирусной репликации на некоторых стадиях могут формироваться стабильные или временные двухцепочечные нуклеиновые кислоты. Ограничение, касающееся однонаправленности переноса информации от ДНК к РНК, не является абсолютным. Оно нарушается ретровирусами, чьи геномы состоят из одноцепочечных молекул РНК. В процессе жизненного цикла РНК ретровирусов переводится в ДНК по механизму обратной транскрипции с участием фермента обратной транскриптазы (РНК-зависимой ДНК-полимеразы). В свою очередь, одноцепочечная ДНК переходит в двухцепочечную форму. Двухцепочечная ДНК становится частью генома клетки-хозяина и наследуется так же, как другие гены. Таким образом, обратная транскрипция позволяет РНК переносить генетическую информацию и предоставлять ее клетке в виде ДНК. Существование механизмов обратной транскрипции и репликации РНК устанавливает важный принцип: информация, содержащаяся в нуклеиновых кислотах любого типа, может быть переведена из одной формы в другую. Чаще всего клетка использует процессы репликации ДНК, транскрипции и трансляции. Однако в редких случаях (вероятно, опосредованных РНКсодержащими вирусами) информация клеточной РНК переводится в ДНК и встраивается обратно в геном. Несмотря на то что обратная транскрипция не играет значительной роли во внутриклеточных процессах, она приобретает значимость при рассмотрении вопроса эволюции генома. Те же принципы сохранения генетической информации справедливы и для больших геномов растений и амфибий, и для сравнительно небольшого генома микоплазмы, и для еще меньших по размеру геномов ДНК- и РНК-содержащих вирусов.
Иерархическая классификация вирусов Эта классификация учитывает такие свойства вирусов как: • морфология (размер, форма, наличие оболочки), • физико-химические свойства (масса, плавучая плотность, pH-, температурная, ионная стабильность), • структура генома (РНК, ДНК, сегментированность, рестрикционная карта, наличие модификаций и т.д.), • свойства макромолекул (набор белков и их функции), • антигенные свойства, • биологические свойства (диапазон хозяев и др.).
Классификация Балтимора Классификация по стратегии репликации вирусов, которая зависит от структуры их геномов. По этому признаку вирусы делят на семь групп.
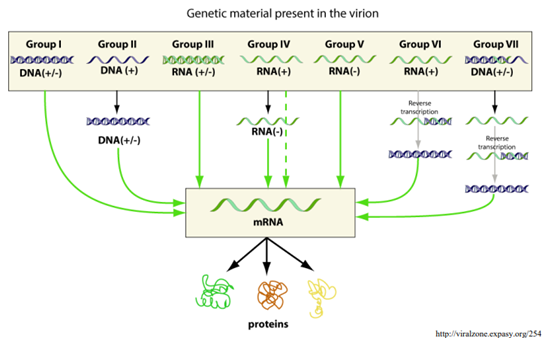
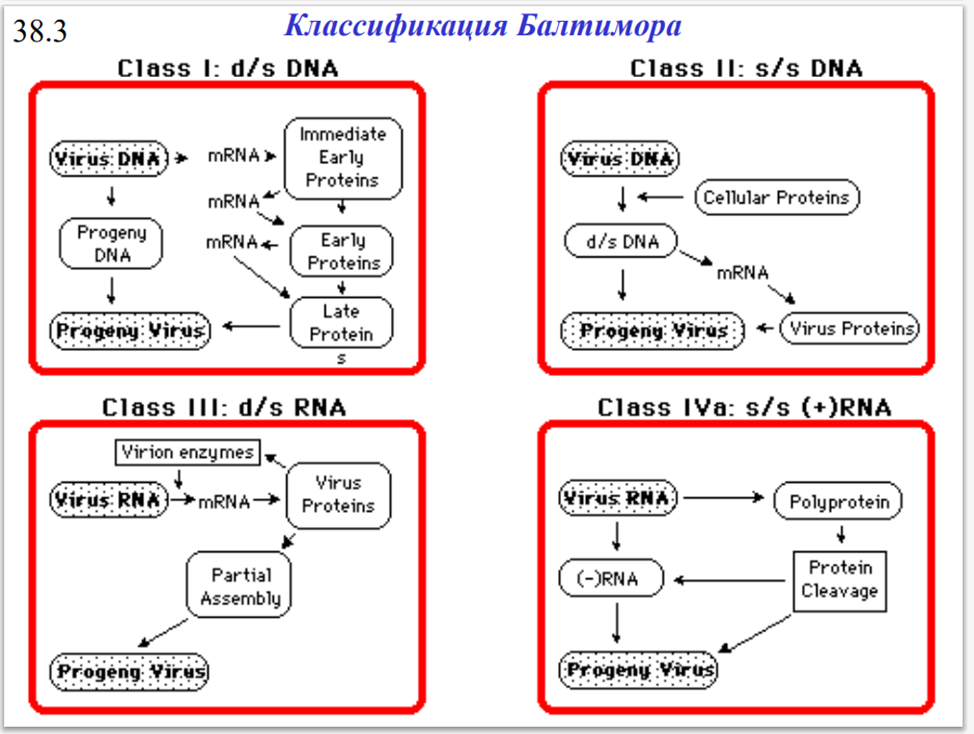

Класс I: Вирусы с двухцепочечной ДНК (dsDNA)
Вирус с двухцепочечной ДНК проникает в ядро клетки-хозяина, прежде чем начать размножаться. Он использует полимеразы клетки-хозяина для репликации своего генома и поэтому сильно зависит от клеточного цикла клетки-хозяина. Таким образом, для размножения вируса клетка должна находиться в процессе репликации. Примерами вирусов I класса являются герпесвирусы, аденовирусы и паповирусы.
Класс II: Вирусы с одноцепочечной ДНК (ssDNA)
Большинство вирусов с одноцепочечной ДНК имеют кольцевые геномы и реплицируются в основном внутри ядра по механизму «катящегося кольца». Примерами вирусов класса II являются Anelloviridae, Circoviridae и Parvoviridae.
Класс III: Вирусы с двухцепочечной РНК (dsRNA)
Вирусы с двухцепочечной РНК реплицируются в сердцевине капсида в цитоплазме клетки-хозяина и в такой же степени зависят от полимераз клетки-хозяина, как и ДНК-вирусы. Геномы вирусов III класса могут быть сегментированы, и, в отличие от вирусов с более сложной трансляцией, каждый ген кодирует только один белок. Примерами вирусов III класса являются реовирусы и бирнавирусы.
Класс IV: Вирусы с одноцепочечной РНК (ssRNA)
Вирусы с ssRNA класса IV имеют РНК-геномы положительного смысла, что означает, что они могут быть непосредственно считаны рибосомами для трансляции в белки. Далее они делятся на вирусы с полицистронной мРНК и вирусы со сложной транскрипцией.
Полицистронная мРНК транслируется в полипротеин, который впоследствии расщепляется на отдельные белки. Вирусы со сложной транскрипцией используют рибосомный сдвиг рамки считывания и протеолитическую обработку для производства нескольких белков из одних и тех же последовательностей генов. Примерами некоторых вирусов IV класса являются Coronaviridae, Flaviviridae, Astroviridae и Picornaviridae.
Класс V: Вирусы с одноцепочечной РНК (ssRNA)
Вирусы класса V имеют геном РНК с негативным смыслом, то есть они должны транскрибироваться вирусной полимеразой для получения считываемой цепи мРНК. Геномы вирусов класса V могут быть сегментированными или несегментированными. Некоторые вирусы из класса V — это Orthomyxoviridae, Paramyxoviridae и Rhabodviridae.
Класс VI: Вирусы с положительной обратной транскриптазой ssRNA
Вирусы группы VI имеют одноцепочечный РНК-геном с положительным смыслом, но реплицируются через промежуточную ДНК. РНК преобразуется в ДНК с помощью обратной транскриптазы, а затем ДНК встраивается в геном хозяина для последующей транскрипции и трансляции с помощью фермента интегразы. Группа VI включает ретровирусы, такие как ВИЧ, а также метавирусы и псевдовирусы.
Класс VII: Вирусы с двухцепочечной ДНК (dsDNA) с обратной транскриптазой
Вирусы класса VII имеют двухцепочечный геном ДНК, но, в отличие от вирусов класса I, они реплицируются с помощью промежуточной одноцепочечной РНК. Двухцепочечный геном ДНК прерывается и впоследствии заполняется, образуя замкнутый круг, служащий матрицей для производства вирусной мРНК. Для воспроизведения генома РНК транскрибируется обратно в ДНК. Вирус гепатита B относится к вирусам класса VII.
Репликация геномов ретровирусов
Геном ретровирусов представлен единственной молекулой одноцепочечной РНК. После проникновения РНК в клетку хозяина вирусный геном подвергается обратной транскрипции, при этом сначала образуется дуплекс РНК- ДНК, а затем двухцепочечная ДНК. Эти этапы предшествуют экспрессии вирусных генов на уровне белков и образованию РНК-геномов.
Фермент, катализирующий комплементарное копирование РНК с образованием ДНК, называется обратной транскриптазой. Он содержится в ретровирусных частицах (вирионах) и активируется после попадания вируса в клетку и разрушения его липид но-гликопротеинов оболочки. Вполне вероятно, что обратной транскрипции способствуют какие-то вспомогательные белки, находящиеся внутри вирионов, поскольку в присутствии последних ферментативная реакция протекает гораздо эффективнее и быстрее, чем в присутствии очищенного фермента.
Обратные транскриптазы ретровирусов по существу являются ДНК-полимеразами, и in vitro могут использовать в качестве матрицы ДНК. Однако гораздо эффективнее они работают, если матрицей является РНК. Как и все ДНК-полимеразы, обратные транскриптазы не способны инициировать синтез новых цепей ДНК. Но если синтез уже инициирован с помощью праймерной РНК или 3-концевого участка ДНК, то фермент эффективно осущест вляет синтез, используя цепь ДНК как матрицу.
Ретровирусы это диплоидные организмы, поскольку каждый вирион содержит две идентичные цепи РНК размером от 8000 до 10000 нуклеотидов. Цепи соединены вблизи своих 5'-концов, однако природа этого нековалентного взаимодействия неизвестна. Области 5 и 3-концов обеих цепей модифицированы, как и у всех эукариотических мРНК (5'-кэпы и 3-полиадениловые хвосты). Рассматривая механизм обратной транскрипции, необходимо отметить наличие пяти структурных элементов у вирусной РНК: 1) прямые повторы на 5 и 3-концах РНК (R); 2) последователь ность из 80-120 нуклеотидов, соседствующая с 5- концевым повтором (U5); 3) последовательность из 170-1200 нуклеотидов, соседствующая с 3-концевым повтором (U3); 4) последовательность из 15-20 нуклеотидов (Р), в пределах которой клеточная тРНК спаривается с ретровирусной РНК, что создает праймер для синтеза первой цепи ДНК; 5) сегмент Ри, находящийся непосредственно перед повтором ИЗ и являющийся сайтом для праймирования второй цепи ДНК; такой сегмент одинаков у РНК всех ретровирусов определенного типа (т.с. его последовательность практически идентична у всех вирусов птиц и отличается от подобной последовательности у вирусов мышей).
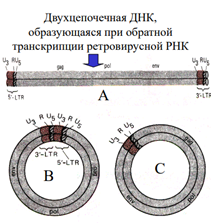
Известны три продукта, образующиеся в результате обратной транскрипции: форма А- линейный дуплекс ДНК с последовательностью U3RUS [длинный концевой повтор LTR (от англ. long terminal repeat)], имеющийся на обоих концах дуплекса; два кольцевых дуплекса ДНК, производных формы А; форма В с LTR-повторами на обоих концах и форма С только с одним LTR. Объяснить образование структур А, В и С при обратной транскрипции вирусной ДНК весьма непросто.
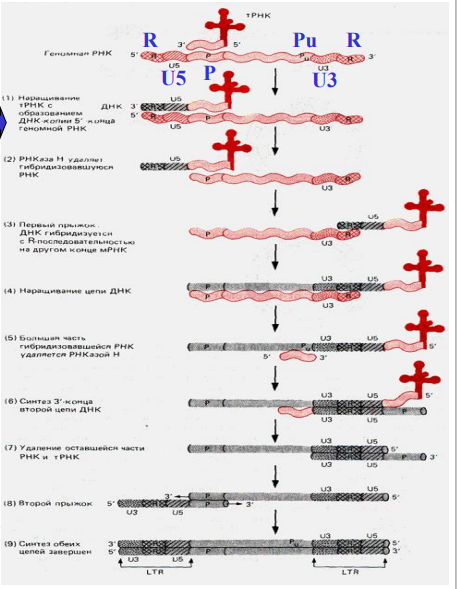
Процесс начинается с наращивания тРНК-праймера на матрицах U5 и R в направле нии 35' (этап 1). Затем РНКаза Н, специфичная к РНК в составе гибридного РНК ДНК-дуплекса, расщепляет сегмент РНК этого дуплекса (этап 2). Поскольку на 3'-конце РНК имеется повтор R, новосинтезированная короткая цепь ДНК «перепрыгивает» на этот конец молекулы мРНК и спаривается там с комплементарным ей участком (этап 3). Далее происходит удлинение цепи ДНК с использованием в качестве матрицы остальной части мРНК (этап 4). К моменту завершения синтеза первой цепи ДНК большая часть вирусной ДНК разрушается РНКазой Н. Затем в предполагаемом сайте связывания праймера (Ри) вблизи повтора U3 инициируется синтез второй цепи ДНК с использованием новой синтезированной первой цепи в качестве матрицы (этап 5). Праймером для синтеза второй цепи может быть РНК, однако как идет синтез второй цепи непрерывно или прерывисто неизвестно. После репликации тРНК-связывающей последовательности на 5'- конце первой цепи ДНК (этап 6) тРНК, по-видимо- му, удаляется (этап 7). Затем новосинтезированная вторая цепь ДНК спаривается с тРНК-связывающей последовательностью первой цепи (этап 8). После удлинения 3-концов каждой цепи завершается образование дуплекса ДНК (этап 9). Обратите внимание, что на каждом конце ДНК-дуплекса имеется прямой повтор последовательности U3RUS-LTR. Кольцевые ДНК по-видимому, образуются либо путем лигирования концов линейной ДНК (Б). либо путем гомологичной рекомбинации между сегментами LTR (B). Удивительно, что такая сложная последовательность реакций протекает без явного участия ферментов репликации клетки-хозяина (геликазы, праймазы, ДНК-связывающего белка, лигазы, топоизомеразы и т.д.).
Репликация двухцепочечной формы ретровирусной ДНК не начинается до тех пор, пока она не встроится в клеточную ДНК. Субстратом для такого интеграционного события является продукт обратной транскрипции линейная дуплексная ДНК.

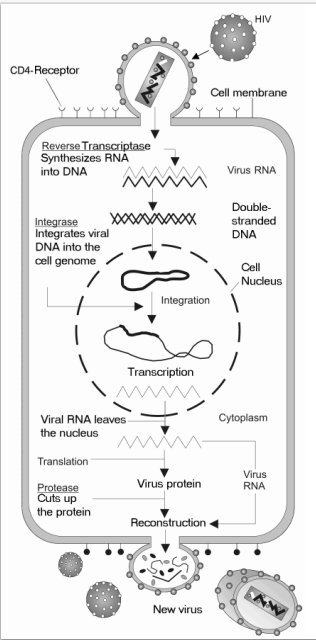
Репликация ВИЧ. Этапы цикла репликации ВИЧ:
• Слияние клетки ВИЧ с поверхностью клетки-хозяина.
• Вход в клетку, РНК ВИЧ, обратная транскриптаза,
интеграза и другие вирусные белки проникают в клетку-хозяина.
• Вирусная ДНК образуется путем обратной транскрипции.
• Вирусная ДНК транспортируется через ядро и
интегрируется в ДНК хозяина.
• Новая вирусная РНК используется в качестве геномной РНК для создания
вирусных белков.
• Новая вирусная РНК и белки перемещаются на поверхность клетки,
и образуется новый, незрелый вирус ВИЧ.
• Созревание вируса и высвобождение протеазой отдельных
белков ВИЧ.