

19. Структурная часть гена эукариот. Мозаичные единицы транскрипции. Экзоны и интроны. Сплайсинг рнк: альтернативный сплайсинг, локализация генов в интронах.
Эукариотический ген – совокупность сегментов ДНК, составляющих экспрессируемую единицу, которая даёт начало одному или нескольким специфическим функциональным продуктам – молекулам РНК либо пептидам.
Основные компоненты структурного гена эукариот:
1. Промотор
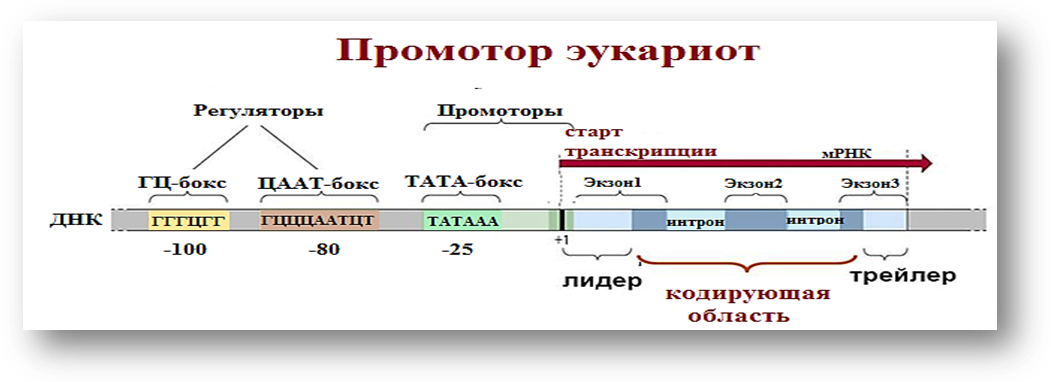
Промотор является ключевым элементом для инициирования транскрипции гена. Это участок ДНК, расположенный перед кодирующей областью гена и необходимый для связывания РНК-полимеразы и других транскрипционных факторов, которые обеспечивают начало синтеза РНК. Промотор не кодирует белок, но играет решающую роль в контроле экспрессии гена.
Структура и функции промотора:
Основной промотор содержит определённые консенсусные последовательности, такие как TATA-box (в случае эукариот), которая находится примерно в 25-30 нуклеотидах перед началом транскрипции. Эта последовательность служит ориентиром для связывания TATA-связывающего белка (TBP), который является частью более крупного комплекса транскрипционных факторов.
В дополнение к TATA-box могут присутствовать другие элементы, такие как CAAT-box и GC-box, которые служат дополнительными регуляторными элементами для усиления или ослабления активности промотора.
Промотор также содержит сайты связывания транскрипционных факторов и может иметь дополнительные элементы, такие как инсилентеры или активаторы, которые регулируют интенсивность транскрипции в зависимости от условий клетки.
2. Экзоны

Экзоны — это участки гена, которые содержат кодирующую информацию. Эти участки в процессе транскрипции переносятся в молекулу РНК, и после сплайсинга они формируют зрелую мРНК, которая затем используется для синтеза белка. Экзоны кодируют аминокислотные последовательности, которые будут составлять белок.
Особенности экзонов:
Экзоны могут быть разделены интронами, которые не кодируют белок и не присутствуют в зрелой РНК.
Экзоны могут включать не только информацию для белка, но и функциональные участки РНК, такие как рРНК и тРНК, которые не преобразуются в белки, но играют важную роль в клеточных процессах (например, в синтезе белков).
3. Интроны
Интроны — это участки генов, которые не кодируют белковую информацию, но присутствуют в первичной РНК. На стадии транскрипции интроны копируются в пРНК, но они удаляются во время сплайсинга, и в зрелой мРНК остаются только экзоны.
Роль и функции интронов:
Регуляторная роль: Хотя интроны не кодируют белок, они могут содержать важные элементы регуляции, которые влияют на экспрессию генов, как в пределах одного гена, так и в межгенном контексте. Это могут быть сайты связывания регуляторных белков или элементы, которые влияют на стабильность РНК.
Альтернативный сплайсинг: Интроны позволяют клетке создавать множество различных вариантов зрелой РНК (альтернативный сплайсинг), что увеличивает разнообразие белков, которые могут быть синтезированы из одного гена. Это важный механизм, способствующий молекулярному разнообразию, а также адаптивной эволюции.
4. Терминатор
Терминатор — это последовательность нуклеотидов, которая сигнализирует о завершении транскрипции. Он распознается РНК-полимеразой и транскрипционными факторами, что приводит к прекращению синтеза РНК и отсоединению РНК-полимеразы от ДНК.
Механизм действия терминации:
В эукариотах существует два основных механизма терминации транскрипции: терминация по типу "петли" (в случае некоторых генной регуляции и с участием определённых белков) и терминация с участием полиаденилирования.
В случае полиаденилирования, который является основным для эукариот, участок ДНК кодирует сигнальную последовательность, которая вызывает добавление поли(A)-хвоста к 3'-концу РНК. Это также способствует завершению транскрипции.
Терминаторный сигнал служит для распознавания точки окончания транскрипции, что критически важно для точности синтеза мРНК и предотвращения "слишком длинных" или ошибочных молекул РНК
Мозаичные единицы транскрипции.
В отличие от прокариотических генов, эукариотические почти всегда коллинеарных своим РНК, многие гены эукариот имеют мозаичное строение. Под мозаичностью подразумевают чередование кодирующих (экзоны) и некодирующих (вставочные последовательности, или интроны) последовательностей в пределах единицы транскрипции. Обычно число интронов на ген возрастает пропорционально длине последовательности, кодирующей белок, а размеры экзонов составляют около 300 п.н. общая длина последовательностей интронов превышает суммарную длину экзонов от 2х до 5ти раз, иногда больше. Интроны располагаются не случайным образом. В генах тРНК они примыкают к петлям антикодонов, а в белок-кодирующих генах часто находятся между сегментами, которые кодируют отдельные структурные или функциональные домены белка.

Альтернативный сплайсинг: множественные белки одного гена.

Альтернативный сплайсинг — это механизм, который позволяет гену продуцировать несколько различных белков или РНК, не изменяя при этом его геномную последовательность. Это важный процесс для эукариот, так как он значительно увеличивает разнообразие молекул, которые могут быть получены из ограниченного числа генов. Основные принципы альтернативного сплайсинга
Во время транскрипции ДНК в про-мРНК происходит вырезание интронов и соединение экзонов. Однако в случае альтернативного сплайсинга экзоны могут быть скомбинированы различными способами, что приводит к образованию различных изоформ зрелой мРНК. Это дает возможность одному гену кодировать несколько функциональных белков или РНК.