

18. Транскрипция у эукариот. Регуляторные участки гена эукариот: инициация транскрипции, энхансеры, сайленсеры и инсуляторы. Терминация транскрипции с участием рнк-полимераз I, II и III.
Механизм транскрипции в ядрах эукариотических клеток намного сложнее, чем у бактерий. Эукариоты имеют три РНК-полимеразы (I, II и III), которые различаются составом комплекса, но имеют некоторые общие субъединицы. Каждая полимераза выполняет особую функцию и присоединяется к специфической промоторной последовательности.
РНК-полимераза I (Pol I) отвечает за синтез только одного типа РНК — прерибосомной РНК (пре-рРНК), которая содержит предшественников 18S, 5,8S и 28S рРНК. Последовательности промоторов Pol I существенно различаются у разных видов. Основная функция РНК-полимеразы II (Pol II) — синтез мРНК и некоторых специализированных РНК. Этот фермент распознает тысячи промоторов с сильно различающимися последовательностями. Многие промоторы Pol II обладают несколькими общими свойствами, в том числе содержат ТАТА-бокс (эукариотическая консенсусная последовательность ТАТА(А/Т)А(А/Т)(А/G)) вблизи позиции –30 и Inr-последовательность (инициатор) вблизи точки начала синтеза РНК в позиции +1. Однако таких промоторов немного, и сложные взаимодействия с регуляторными белками определяют функцию Pol II на многих промоторах, у которых эти особенности отсутствуют.
РНК-полимераза III (Pol III) образует молекулы тРНК, 5S рРНК и некоторые другие небольшие специализированные РНК. Промоторы, распознаваемые Pol III, хорошо охарактеризованы. Интересно, что некоторые последовательности, необходимые Pol III для осуществления регулируемой инициации транскрипции, расположены в самом гене, а другие локализованы более традиционно — выше точки начала транскрипции.
РНК-полимераза II играет ведущую роль в экспрессии генов эукариот и поэтому активно изучалась. Фермент Pol II, выделенный из дрожжей или клеток человека, состоит из 12 субъединиц с совокупной молекулярной массой 510 000. Самая крупная субъединица RBP1 во многом гомоло- гична β’-субъединице бактериальной РНК-полимеразы. Другая субъединица RBP2 по структуре похожа на бактериальную -субъединицу, еще две субъединицы RBP3 и RP11 имеют некоторую структурную гомологию с двумя бактериальными α-субъединицами. Полимеразе Pol II приходится действовать в гораздо более сложных геномах и взаимодействовать с намного более сложно упакованными молекулами ДНК, чем у бактерий. Дополнительная сложность полимеразы эукариот объясняется необходимостью взаимодействовать со множеством белковых факторов.
Самая крупная субъединица Pol II (RBP1) обладает необычным свойством: на C-конце она имеет длинную аминокислотную последовательность, состоящую из многократно повторяющейся консенсусной последовательности семи аминокислот –YSPTSPS–. В ферменте дрожжей содержится 27 таких повторов (18 из них в точности соответствуют консенсусной последовательности), а в ферментах мыши и человека — по 52 повтора (21 точный). Этот С-концевой домен CTD (от англ. carboxyl-terminal domain) отделен от основного тела фермента неструктурированной линкерной последовательностью. Как показано ниже, CTD необходим для выполнения множества функций Pol II.
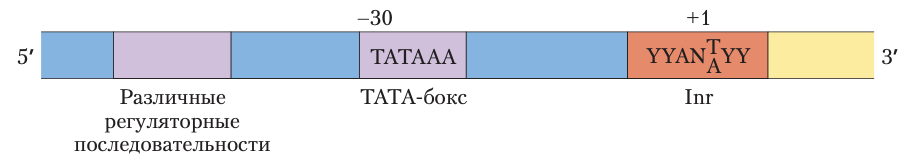
Последовательности промотора, распознаваемые эукариотической РНК-полимеразой II.
Точкой сборки белков преинициаторных комплексов Pol II является ТАТА-бокс. ДНК в инициаторной последовательности (Inr) расплетается, а точка начала транскрипции обычно находится в этой последовательности или в непосред- ственной близости от нее. В представленной здесь консенсусной последовательности Inr N обозначает любой нуклеотид, а Y — пиримидиновый нуклеотид. Многие дополнительные последовательности служат участками связывания для большого числа белков, влияющих на активность Pol II. Эти последовательности важны для регуляции промоторов Pol II и значительно различаются по типу и количеству. Многие последовательности расположены на расстоянии нескольких сотен пар оснований от ТАТА-бокса с 5’-стороны, другие могут быть удалены на тысячи пар оснований.Во многих промоторах Pol II отсутствует либо ТАТА-бокс, либо элемент Inr, либо обе последовательности. Один или несколько транскрипционных факторов распознают дополнительные последовательности около ТАТА-бокса или ниже по последовательности (на рисунке справа).
Для формирования активного транскрипционного комплекса РНК-полимераза II нуждается в ряде других белков, называемых факторами транскрипции. Основные факторы транскрипции для каждого промотора Pol II (обычно обозначаются TFII с дополнительной идентификацией) достаточно консервативны у всех эукариот. В процессе транскрипции под действием фермента Pol II можно выделить несколько стадий: сборку, инициацию, элонгацию, терминацию; на каждой стадии участвуют определенные белки.
Сборка РНК-полимеразы и факторов транскрипции на промоторе Образование закрытого комплекса начинается с взаимодействия ТАТА-связывающего белка (ТВР) с ТАТА- боксом (стадия 1). В промоторы, лишенные ТАТА-бокса, TBP поступает как часть мультисубъединичного комплекса (TFIID); элементы последовательности, которые управляют связыванием TFIID с этими промоторами, не содержащими TATA, изучены плохо. ТВР, в свою очередь, связывается с фактором транскрипции TFIIB, затем связывается TFIIA и вместе с TFIIB помогает стабилизировать комплекс TBP–ДНК. TFIIB обеспечивает важную связь с ДНК-полимеразой II, и комплекс TFIIB–TBP соединяется с другим комплексом, состоящим из TFIIF и Pol II. Фактор TFIIF помогает точной стыковке Pol II с промотором, взаимодействуя с TFIIB и одновременно ослабляя связывание полимеразы с неспецифическими участками ДНК. Наконец, присоединяются TFIIE и TFIIH — образуется закрытый комплекс. Фактор TFIIH состоит из нескольких субъединиц и обладает ДНК-хеликазной активностью, за счет которой начинается раскручивание ДНК вблизи точки начала транскрипции РНК (процесс нужда- ется в гидролизе ATP) и образуется открытый комплекс (стадия 2 на рис. ниже). С учетом всех субъединиц разных факторов (включая TFIIA и субъединицы TFIID) этот активный инициирующий комплекс может содержать более 50 полипептидов.
Инициация цепи РНК и высвобождение промотора
На стадии инициации TFIIH выполняет дополнительную функцию. Киназная активность одной из его субъединиц фосфорилирует последовательность CTD в Pol II во многих местах. Несколько других протеинкиназ, включая CDK9 (циклинзависимая киназа 9), которая является частью комплекса pTEFb (от англ. positive transcription elongation factor b — положительный фактор транскрипции и элонгации b), также фосфорилируют CTD, особенно остатки серина. Это приводит к конформационным изменениям всего комплекса и инициирует транскрипцию. Фосфорилирование CTD также важно для следующей стадии элонгации, причем степень фосфорилирования CTD изменяется по мере продвижения транскрипции. Эти изменения влияют на взаимодействия транскрипционного комплекса с другими белками и ферментами, так что при инициации транскрипции в комплексе связаны другие белки, нежели на более поздних стадиях. Некоторые из этих белков участвуют в процессинге транскрипта.
В процессе синтеза первых 60–70 нуклеотидов РНК сначала из комплекса выходит TFIIE, потом TFIIH, и Pol II начинает стадию элонгации.
Элонгация, терминация и высвобождение
На протяжении всей элонгации TFIIF остается связанным с Pol II. На этой стадии активность полимеразы значительно усиливается белками, называемыми факторами элонгации. Факторы элонгации, некоторые из которых связаны с фосфорилированным CTD, препятствуют остановке транскрипции, а также координируют взаимодействия между белковыми комплексами, вовлеченными в посттранскрипционный процессинг молекул мРНК. Как только синтез транскрипта РНК завершается, транскрипция прекращается. Полимераза II дефосфорилируется, после чего она готова инициировать транскрипцию другого участка (стадии 3 и 5 на рис. ниже).
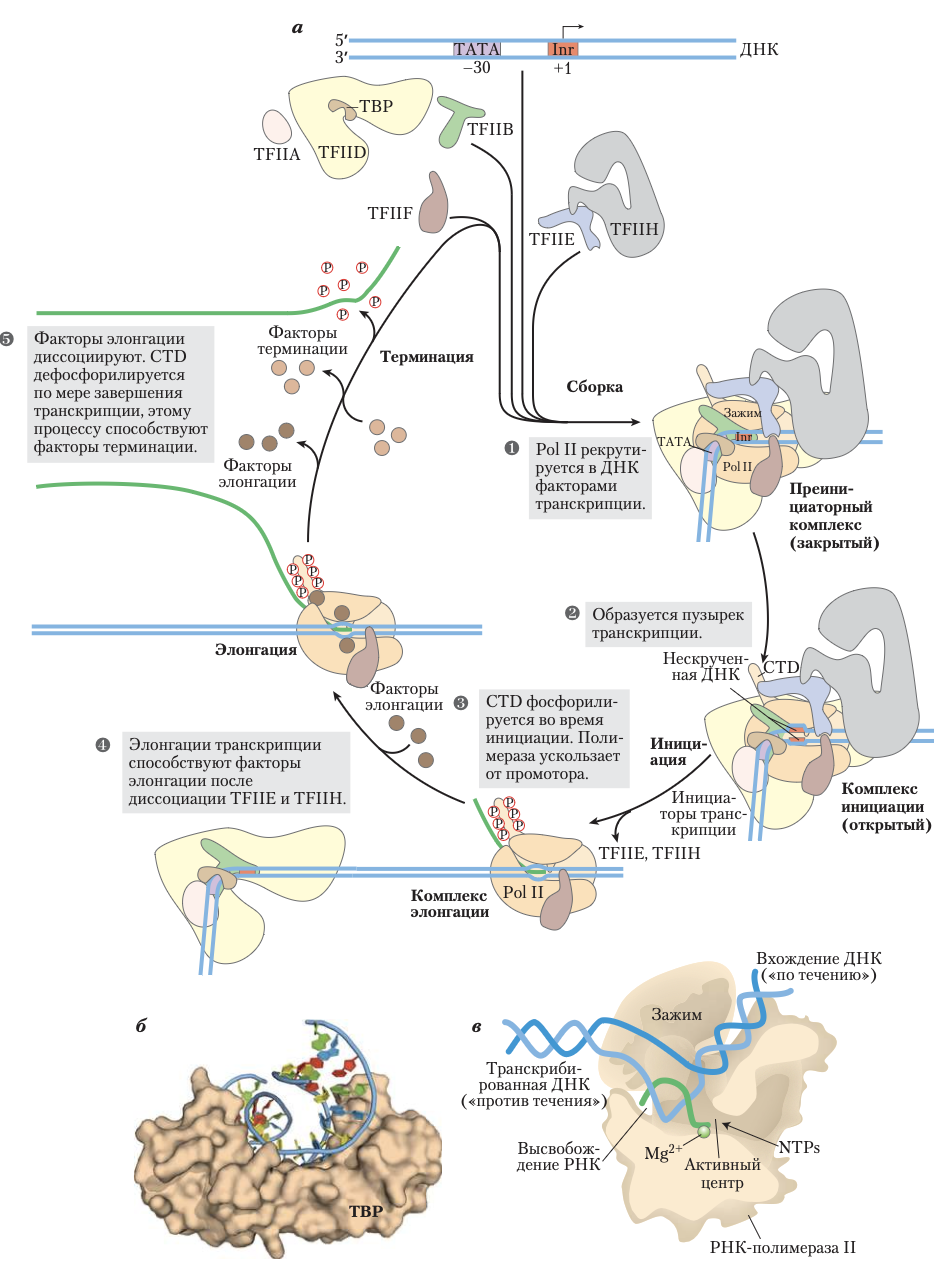
На рисунке выше показана схема транскрипции с промоторов РНК-полимеразы II. а) ТВР (часто с TFIIA, а иногда с TFIID) и TFIIB последовательно связываются с промотором. Затем к комплексу присоединяются TFIIF и Pol II. Дальнейшее присоединение TFIIE и TFIIH приводит к формированию закрытого комплекса. Внутри комплекса происходит раскручивание ДНК в области Inr за счет активности TFIIH-хеликазы и, возможно, TFIIE, что приводит к открытию комплекса, — сборка завершается. С-концевой домен самой крупной субъединицы Pol II фосфорилируется под действием TFIIH, затем полимераза оставляет промотор и инициирует транскрипцию. Элонгация сопровождается высвобождением многих факторов транскрипции и усиливается факторами элонгации. После терминации Pol II высвобождается, дефосфорилируется и может участвовать в новом синтезе. б) ТВР человека, связанный с ДНК. В этом комплексе ДНК изгибается, открывая малую бороздку и обеспечивая образование характерных водородных связей между белком и ДНК. в) Схема (в разрезе) элонгации транскрипции, катализируемой кор-ферментом Pol II.
Эукариотические РНК-полимеразы имеют небольшое сродство к своим промоторам (или не имеют его совсем), поэтому инициация транскрипции почти всегда зависит от действия многочисленных белков-активаторов.
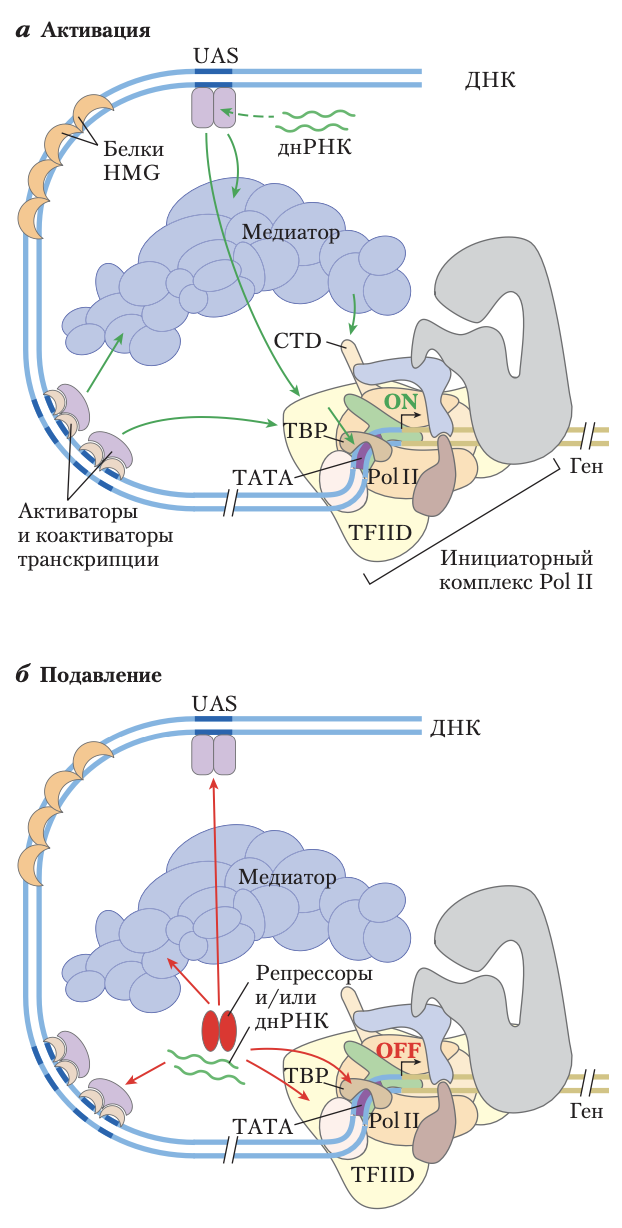
Эукариотические промоторы и регуляторные белки
РНК-полимераза II и ассоциированные с ней основные факторы транскрипции образуют преинициаторный комплекс на последовательностях ТАТА-бокса и Inr в соответствующем промоторе; этому процессу способствуют активаторы транскрипции, действующие через коактиваторы (медиатор, TFIID или оба). a) Составной промотор с элементами типичных последовательностей и белковыми комплексами, существующими как в дрожжах, так и в высших эукариотах. С-концевой домен (CTD) Pol II — важный участок взаимодействия с медиатором и другими белковыми комплексами. Ферменты модификации гистонов (не показаны) катализируют метилирование и ацетилирование; ферменты ремоделирования изменяют состав и расположение нуклеосом. Активаторы транскрипции имеют различимые ДНК-связывающие домены и активационные домены. В некоторых случаях на их функцию оказывает влияние взаимодействие с длинными некодирующими РНК (днРНК). Зелеными стрелками показаны типичные взаимодействия, часто необходимые для активации транскрипции, как обсуждается в тексте. Белки HMG — распространенный тип архитектурных регуляторных белков, способствующих образованию петель ДНК для сближения компонентов системы, связанных в отдаленных друг от друга участках. б) Репрессоры транскрипции эукариот действуют посредством нескольких механизмов. Некоторые напрямую связываются с ДНК, вытесняя необходимый для активации белковый комплекс (не показан); другие предотвращают активацию, взаимодействуя с различными частями транскрипционных или активаторных комплексов. Возможные участки взаимодействия показаны красными стрелками.
Только в пределах участка между двумя инсуляторами энхансерные последовательности, связавшись с белками-активаторами, могут образовать петлю и осуществить взаимодействие с промотором
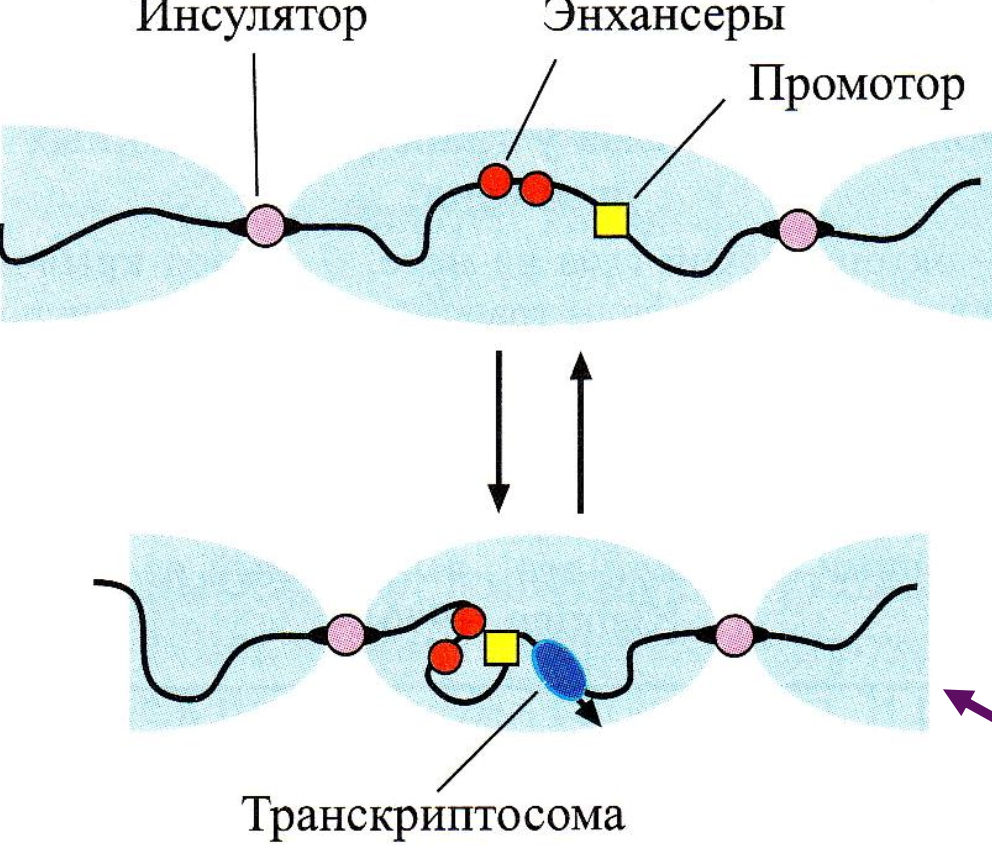
Механизмы, которые определяют 3'-конец соответствующих функциональных РНК-продуктов, уникальны для каждой полимеразной системы.
Сигналом к терминации транскрипции с участием Pol I служат высококонсервативные последовательности значащей цепи (у мышей длиной 18 п.н. 5'-AGGTCGACCAG(TA/AT)NTCCG-3', так называемый SalI-бокс), которой обычно предшествует один или несколько пиримидиновых кластеров. Tерминация в этих сайтах происходит при связывании специфических белков с
последовательностями ДНК.
3'-концы РНК, синтезированных при помощи РНК-полимеразы III, как правило, соответствуют сайтам, в которых заканчивается транскрипция. Для терминации транскрипции нужен только сам фермент и кластер дезоксиаденилатных остатков на матричной цепи и определенные фланкирующие его последовательности.
Транскрипция Pol II многих генов белков эукариот осуществляется во множественных сайтах последовательности ДНК протяженностью в сотни, а иногда и тысячи пар нуклеотидов.
Некоторые гены белков, напротив, имеют четко определенные сигналы терминации транскрипции. Транскрипция заканчивается перед сигнальной последовательностью, в каком бы месте ДНК она ни находилась. Этот сигнал напоминают ро-зависимые сигналы терминации у Е. coli. Терминация транскрипции связана с полиаденилированием. Последовательность ААТААА - обязательный элемент сигнала полиаденелирования, участвует также в терминации транскрипции.