

Филогенез пищеварительной системы
.pdfТЕМАТИЧЕСКИЙ БЛОК №19
Тема: Филогенез систем органов животных и человека. Пищеварительная система.
Цель: Получить представление об эволюции процесса питания и способах переваривания пищи. Знать основные этапы развития пищеварительной системы у беспозвоночных животных.
Уметь охарактеризовать направления эволюции пищеварительной системы позвоночных.
Получить представление о филонтогенетических пороках развития пищеварительной системы у человека.
Основные вопросы темы:
1.Этапы эволюции процесса питания и способов переваривания пищи.
2.Основные направления эволюционных преобразований пищеварительной системы у беспозвоночных.
3.Основные направления эволюционных преобразований пищеварительной системы у
позвоночных.
4.Понятие о филонтогенетических пороках развития пищеварительной системы у
человека.
Содержание темы
Общая характеристика пищеварительной системы
Пищеварительная система представляет собой пищеварительный тракт, образованный желудком, тонким и толстым кишечником, а также примыкающими к нему железистыми образованиями (слюнные железы, печень с желчным пузырем, поджелудочная железа).
Пищеварительная система выполняет целый ряд функций. Непосредственное отношение к процессу пищеварения имеют:
1. Сенсорная функция обеспечивается сенсорными рецепторами пищеварительного тракта, и заключается в восприятии физико-химических параметров принимаемой пищи
(температуры, вкусовых качеств, консистенции и др.), передаче информации в ЦНС для се оценки.
Во всех отделах пищеварительного тракта имеются механореценторы, реагирующие на моторику,
тонус, степень растяжения стенок желудка, кишечника, протоков желез, многочисленных сфинктеров. Их сигналы используются для координации моторики органов пищеварительного тракта.
2. Двигательная или моторная функция, осуществляется за счет мускулатуры пищеварительного аппарата и включает в себя процессы измельчения пищи в полости рта,
глотания, перемещения пищевого комка, а также удалении непереваренных остатков.
3. Секреторная функция связана с выработкой железистыми клетками пищеварительных секретов: слюны, желудочного сока, сока поджелудочной железы, желчи,
способных, благодаря наличию в их составе ферментов, расщеплять белки, жиры и углеводы на простые химические соединения.
4. Всасывательная функция подразумевает проникновение через стенку пищеварительного тракта в кровь и лимфу продуктов гидролитического расщепления пищи:
аминокислот, моносахаров, жирных кислот и глицерина, а также воды, электролитов, витаминов и пр.
Кроме вышеназванных, пищеварительная система выполняет также следующие функции:
5. Защитная функция обеспечивается неспецифическими и специфическими механизмами. Неспецифическая защита реализуется за счет: рефлекторного отторжения
(выплевывание или рвота) некачественной пищи (имеющей нарушения органолептических свойств, содержащей механические примеси и пр.); бактерицидного и бактериостатического действия компонентов пищеварительных соков и др. Специфическая система защиты представляет собой совокупность структур и элементов лимфоидной ткани, расположенных на протяжении всего пищеварительного тракта и являющихся частью иммунной системы организма:
лимфоидные фолликулы (пейеровы бляшки тонкой кишки, миндалины глоточного кольца), Т-
лимфоидные клетки и плазматические элементы слизистой оболочки, а также малые лимфоидные клетки. Иммунокомпетентные клетки, локализующиеся в пищеварительном тракте, осуществляют распознание антигенов химуса, обеспечивают местный иммунный ответ и формируют иммунную память.
6.Инкреторная функция определяется выработкой гормонов (гастрин, секретин,
холецистокинин-панкреозимин и др.), оказывающих влияние на моторную и секреторную функции системы пищеварения, а также на интенсивность некоторых метаболических процессов
7. Экскреторная функция представляет собой совокупность процессов, благодаря которым вещества выделяются из внутренней среды организма в просвет пищеварительного канала с последующим выведением из организма. Экскретируемыми веществами могут быть продукты обмена аммиака, мочевины, соли тяжелых металлов и пр.
Эволюция процесса питания и способов переваривания пищи
Питание – это процесс приобретения веществ, необходимых для покрытия энергетических и пластических потребностей организма. В процессе эволюции возникли два основных способа питания: автотрофное и гетеротрофное. При автотрофном питании организмы способны создавать органические вещества из неорганических – углекислого газа, воды, минеральных солей. Такой способ питания характерен для представителей царства растений и некоторых бактерий. Для осуществления процессов синтеза органических веществ из неорганических необходима энергия. Автотрофные организмы могут синтезировать органические вещества за счет
энергии солнечного света. Такие организмы называются фототрофами (практически все растения,
зеленые протисты, цианобактерии). Организмы, которые для осуществления синтеза органических веществ используют энергию окисления некоторых химических веществ, называются хемотрофами (железобактерии, бесцветные серобактерии, нитрифицирующие бактерии).
При гетеротрофном питании организмы нуждаются в поступлении готовых питательных веществ из вне. К числу последних относятся представители царства животных, царства грибов и большинство бактерий. Существует несколько типов гетеротрофного питания: голозойный,
сапрофитный, симбиотический и паразитический.
При голозойном типе питания организмы обладают специальным пищеварительным трактом, где захваченная внутрь тела пища, подвергается перевариванию и всасыванию. Такой тип питания характерен для большинства многоклеточных животных, а также свойственен и некоторым одноклеточным (например, амебе), осуществляющим фагоцитоз и пищеварение в
фаголизосомах.
Сапрофитный тип питания характерен для организмов, питающихся мертвым или разлагающимся органическим материалом. Все сапрофитные организмы выделяют ферменты непосредственно на потенциальный продукт питания, который под воздействием этих ферментов подвергается перевариванию. Растворимые конечные продукты такого переваривания
всасываются и ассимилируются сапрофитом. К сапрофитам относятся грибы и многие бактерии.
Симбиотический тип питания характерен для симбиотических организмов и может реализовываться в форме мутуализма или комменсализма. В первом случае отношения между симбионтами являются взаимовыгодными, во втором – один из симбионтов извлекает пользу из
симбиотических взаимоотношений, тогда как его партнер не получает ни пользы, ни вреда.
При паразитическом типе питания организмы (паразиты) получают органические вещества от организма-хозяина, тем самым причиняя ему вред. Паразитический способ питания характерен
для некоторых бактерий, протистов, животных и некоторых высших растений.
Способы добывания и поглощения пищи у гетеротрофных организмов весьма разнообразны, однако пути превращения пищи в удобную для усвоения форму у многих организмов сходны и состоят из следующих этапов: заглатывание, переваривание, всасывание,
усвоение и выделение (экскреция). С точки зрения эволюционных преобразований наиболее
сложным из перечисленных этапов является этап переваривания.
Переваривание (пищеварение) – расщепление крупных органических молекул на компоненты, лишенные видовой специфичности и пригодные к всасыванию и участию в обмене
веществ. Переваривание можно разделить на два этапа: механическое и химическое
(ферментативное). Расщепление сложных молекул происходит с присоединением к ним молекулы воды, поэтому реакции, осуществляющие химическое переваривание, называются

гидролитическими. Существует три основных типа переваривания (пищеварения): внеклеточное
(дистантное), внутриклеточное, и мембранное.
Рис. 1. Локализация гидролиза пищевых веществ при различных типах пищеварения1:
А— внеклеточное (дистантное); Б — внутриклеточное, В — мембранное пищеварение; 1
—внеклеточная жидкость; 2 — внутриклеточная жидкость; 3 — внутриклеточная вакуоль; 4 —
ядро; 5 — клеточная мембрана; 6 — ферменты.
Внеклеточное (дистантное) пищеварение характеризуется тем, что синтезированные секреторными клетками ферменты выделяются во внеклеточную среду, где реализуется их гидролитический эффект. Данный тип пищеварения представляет собой начальный этап химической обработки пищи. Впервые секреция гидролитических ферментов из клеток в пищеварительные полости появилась у кишечнополостных. У кольчатых червей, ракообразных,
насекомых, головоногих и хордовых внеклеточное пищеварение преобладает. У большинства высокоорганизованных животных секреторные клетки расположены достаточно далеко от пищеварительных полостей, где реализуется действие гидролаз, поэтому внеклеточное пищеварение определяется как дистантное. Если дистантное пищеварение происходит в специальных полостях, то оно определяется как полостное. Дистантное пищеварение может происходить и за пределами организма, в этом случае пищеварение является дистантным внеполостным.
Внутриклеточное пищеварение. Термин используется для обозначения всех случаев, когда нерасщепленные или частично расщепленные пищевые субстраты проникают внутрь клетки, где подвергаются гидролизу ферментами цитоплазмы. Внутриклеточное пищеварение может происходить не только в цитоплазме, но и в специальных внутриклеточных полостях – пищеварительных вакуолях, образующихся при пиноили фагоцитозе при участии гидролитических ферментов лизосом. Внутриклеточное пищеварение характерно для
1 БСЭ. Третье издание – М.:, 1975, -С 1756
одноклеточных и низших многоклеточных организмов. У высших животных примером внутриклеточного пищеварения могут служить фагоцитарные свойства лейкоцитов и клеток ретикуло-эндотелиальной системы.
Мембранное пищеварение осуществляется ферментами, локализованными на структурах клеточной мембраны. У человека и высших животных мембранное пищеварение происходит на поверхности мембран микроворсинок кишечных клеток и является механизмом промежуточных и заключительных стадий гидролиза. Данный вид пищеварения обеспечивает сопряжение пищеварительных и транспортных процессов, что достигается благодаря специальной организации пищеварительных и транспортных функций клеточной мембраны в виде своеобразного пищеварительно-транспортного «конвейера», способствующего передаче конечных продуктов гидролиза с фермента на переносчика и предотвращает конкуренцию между ними за вход в транспортную систему мембраны.
Считается, что внутриклеточное пищеварение является эволюционно самым древним типом, так как оно распространено у простейших и наиболее примитивных многоклеточных организмов. Возникновение внеклеточного пищеварения в процессе эволюция происходило не одновременно для всех категорий пищи у одного и того же животного, и поэтому оно претерпело немало изменений. В частности, дальнейшее усовершенствование пищеварения для наиболее высокоорганизованных организмов пошло по пути развития мембранного пищеварения наряду с внеклеточным. Сочетание увеличения суммарной площади, на которой происходит гидролиз пищевых веществ при мембранном пищеварении, с эффективной работой ферментов при внеклеточном пищеварении обеспечивает нутритивные потребности высокоорганизованных организмов.
Разнообразие типов гетеротрофного питания, уровень организации организмов – гетеротрофов, на первый взгляд может свидетельствовать о более позднем, по сравнению с автотрофным типом, возникновении данного типа питания в процессе эволюции. В свете современных данных складывается представление о том, что фотосинтезирующая автотрофная биосфера, была образованием вторичным и возникла на основе биосферы иного биогеохимического типа, из чего следует вывод первичности гетеротрофной формы обмена веществ в первичных организмах. Существенным аргументом такого вывода является тот факт,
что у гетеротрофных организмов не встречается никаких признаков или рудиментарных остатков тех специфических ферментных комплексов и биохимических реакций, которые необходимы для автотрофного способа питания. Скорее всего первичная биосфера нашей планеты была насыщена гетеротрофами, но этот период был коротким, потому что запасы органических веществ не могли возобновляться так же, как запасы автотрофных организмов. Первичные гетеротрофы,
размножаясь, исчерпали свою питательную базу. Поэтому, достигнув максимальной биомассы,
они должны были вымирать или перейти к автотрофному фотосинтетическому способу питания.
Однако первичная поверхность новорожденной Земли, лишенная свободного кислорода,
облучалась ультрафиолетовой радиацией Солнца. Поэтому, можно допустить, что первичные фотохимические механизмы, принимавшие участие в последовательном синтезе органических веществ, а позже и живых организмов, первоначально использовали радиацию в ультрафиолетовой области спектра. Только после возникновения озонового экрана в связи с появлением свободного кислорода как побочного продукта фотосинтеза автотрофный фотосинтетический процесс начал использовать излучение в видимой части солнечного спектра.
Эволюция пищеварительной системы беспозвоночных
Развитие пищеварительной системы беспозвоночных идёт в направлении появления, на основе слепо замкнутой гастроваскулярной полости, пищеварительной трубки, ее удлинения и дифференцировки; появления ротового аппарата и его усложнения; появления и дифференцировки пищеварительных желёз.
Простейшие (Protozoa). У представителей подцарства Protozoa – простейших - организм представлен одной клеткой. Для них характерны три способа питания: автотрофный,
гетеротрофный и миксотрофный. Два последних типа питания представлены только у части видов жгутиконосцев. Все остальные простейшие являются гетеротрофами. Вероятно, автотрофный тип питания у одноклеточных в процессе эволюции никогда не реализовывался в чистом виде, так как все известные одноклеточные, имеющие хлоропласты, используют для питания также и растворенные органические вещества. Гетеротрофный способ питания у простейших может осуществляться тремя способами: за счет мембранного транспорта, пиноцитоза и фагоцитоза.
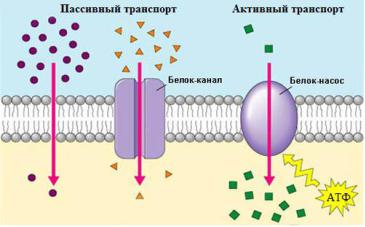
Мембранный транспорт у простейших реализуется путем диффузии (пассивный транспорт) и активного транспорта. Каналы мембраны обеспечивают диффузию воды, ионов и небольших органических молекул по градиенту концентрации, однако некоторые мембранные белки способны активно переносить молекулы и ионы внутрь клетки или наружу против градиента концентрации (рис. 2).
Рис. 2. Мембранный транспорт
Пиноцитоз заключается в захвате клеточной поверхностью жидкости с растворенными в ней веществами. При этом на поверхности плазмалеммы образуется небольшая ямка, которая

затем отшнуровывается в виде небольшого пузырька. При более сложном варианте пиноцитоза ямка на поверхности плазмалеммы углубляется, так что формируется тонкий канал. От дна этого канала происходит отшнуровывание мелких пузырьков. У некоторых простейших такие каналы являются временными образованиями, но у части других простейших они имеют постоянное местоположение, в этом случае начальная часть такого канала называется ультрацитостомом (рис.
3).
Фагоцитоз представляет собой захватывание крупных оформленных частиц.
Формирование фагоцитарных вакуолей может происходить с участием псевдоподий,
охватывающих пищевую частичку или при изменении профиля клеточной мембраны – прогибания ее вовнутрь. В результате поглощенная пищевая частица оказывается внутри пищеварительной вакуоли, окруженной мембраной. Переваривание пищи происходит за счет пищеварительных ферментов, выделяемых лизосомами и поступающих внутрь вакуоли. Продукты внутриклеточного пищеварения поступают в цитоплазму клетки простейшего, где в последующем используются для удовлетворения энергетических потребностей организма или участия в реакциях биосинтеза. Непереваренные остатки выбрасываются во внешнюю среду путем экзоцитоза. У некоторых простейших (амебы) фагоцитарные вакуоли могут формироваться на любом участке тела при этом отсутствуют специальные органеллы, обеспечивающие прием пищи отсутствуют. У других представителей простейших (инфузории) имеются такие специальные органеллы (цитостом, цитофаринкс и др.), имеющие определенную локализацию. Удаление непереваренных остатков может происходить путем экзоцитоза через любой участок поверхности клетки (амебы) или на определенном участке через порошицу, или цитопрокт (инфузории). В
любом из этих случаев механизм экзоцитоза сводится к слиянию мембраны вакуоли и поверхностной мембраны клетки (рис. 3).
Рис. 3. Пиноцитоз и фагоцитоз
Радиально-симметричные беспозвоночные
Кишечнополостные (Coelenterata) – двухслойные животные с радиальной симметрией
(полипы и медузы). Тело кишечополостных образовано двумя слоями клеток – наружным
(эктодермой) и внутренним (энтодермой). Важным эволюционным приобретением кишечнополостных является появление слепо замкнутой гастроваскулярной полости. С внешней средой полость соединена ротовым отверстием, которое служит и для приема пищи и для выбрасывания непереваренных остатков. Изнутри полость выстлана энтодермой, железистые клетки, которой выделяют в нее пищеварительные ферменты. В гастроваскулярной полости происходит частичное переваривание добычи, поступающей в нее через рот. Затем кусочки пищи захватываются клетками энтодермы, и переваривание завершается в пищеварительных вакуолях.
Таким образом, переваривание у этих животных частично внеклеточное, а частично внутриклеточное. Внеклеточное пищеварение обеспечивает кишечнополостным возможность поедать крупную добычу и питаться лишь время от времени. У гидроидных полипов пищеварительная система представляет собой цельный гладкостенный мешок. У других форм кишечнополостных наблюдаются различные приспособления к увеличению поверхности гастродермы. Так у сцифоидных и коралловых полипов увеличение поверхности гастродермы происходит благодаря наличию септ, представляющих собой отходящие от стенки тела неполные перегородки. У медуз формируется от 4 (гидромедузы) до 16 (сцифомедузы) радиальных каналов,
отходящих от мешковидной части пищеварительной системы и впадающих в кольцевой канал,
идущий по краю зонтика. Весь комплекс каналов образует так называемую гастроваскулярную систему. Такой тип пищеварительной системы называется гастроваскулярным. Третьим вариантом увеличения поверхности гастродермы является формирование многочисленных тонких выростов,
гастральных нитей, характерных для сцифоидных медуз. Важную роль в процессе питания играет наличие щупалец, расположенных вокруг ротового отверстия, и стрекательных клеток. Последние оглушают или убивают добычу, которая затем загоняется в полость тела с помощью щупалец.
Большинство кишечнополостных являются макрофагами, так как способны поглощать пищу в виде крупных частиц. Но есть и такие представители типа, которые, напротив, поглощают пищу в виде мелких частиц, являясь микрофагами, или такие кишечнополостные, которые способны питаться и как макрофаги, и как микрофаги. К числу последних относят, например,
часть коралловых полипов. Большинство кишечнополостных являются хищниками, питающимися живыми объектами. При этом, предварительно убитая или оглушенная с помощью стрекательных клеток, добыча подносится щупальцами к ротовому отверстию и заглатывается. Некоторые виды кишечнополостных питаются детритом (совокупность взвешенных в воде и осевших на дно водоема органо-минеральных частиц биогенного и абиогенного происхождения). При питании детритом вода, несущая в себе пищевые частицы, загоняется в гастральную полость биением ресничек. Некоторые виды используют для сбора пищи слизь. Некоторые из кишечнополостных ведут колониальный образ жизни (гидроидные полипы). Соединенные между собой с помощью

полых трубочек гидранты, имеют общую пищеварительную полость – энтерон. Колониальная организация обеспечивает высокую эффективность питания.
Процесс пищеварения кишечнополостных можно рассмотреть на примере гидры (рис. 4 А).
Гидра питается мелкими беспозвоночными. Добыча захватывается щупальцами с помощью стрекательных клеток, яд которых быстро парализует мелких жертв. Координированными движениями щупалец добыча подносится ко рту, а затем с помощью сокращений тела гидра
«надевается» на жертву. В состав энтодермы, выстилающей кишечную полость гидры изнутри,
входят пищеварительно-мускульные и железистые клетки (рис. 4 Б). Пищеварение начинается в кишечной полости (полостное пищеварение), куда железистыми клетками выделяется пищеварительный сок, который разжижает и частично переваривает пищу. Далее пищеварение осуществляется внутри пищеварительных вакуолей (внутриклеточное пищеварение). Каждая из пищеварительно-мускульных клеток энтодермы имеет 1-3 жгутика. Колеблющиеся жгутики создают ток воды, которым пищевые частички подгоняются к клеткам. Кроме того пищеварительно-мускульные клетки способны образовывать ложноножки, с помощью которых мелкая пища захватывается, а затем переваривается в пищеварительных вакуолях. Если в пищеварительной полости оказываются остатки жертвы, которые невозможно переварить, и
отходы клеточного обмена, гидра сжимается и опорожняется через то же отверстие в полости,
через которое пища поступает.
Рис. 4. Пищеварение гидры:
А – общее строение гидры; Б – элементы экто- и энтодермы гидры
Билатерально-паренхиматозные беспозвоночные
Плоские черви (Рlathelminthes) – трехслойные животные, в процессе онтогенеза которых все ткани и органы развиваются из трех зародышевых листков: эктодермы, энтодермы и мезодермы. Полость тела отсутствует. Внутреннее пространство тела заполнено паренхимой.
Пищеварительная система представлена двумя отделами: эктодермальным передним (глотка и

пищевод) и энтодермальным средним, замкнутым слепо. Отделы функционально разделены: через переднюю кишку происходит поглощение пищи, а в средней кишке она расщепляется, и нужные вещества всасываются в тканевую жидкость паренхимы. Пищеварение внутриполостное и внутриклеточное. Пищеварительные ферменты выделяют железистые клетки, расположенные в среднем отделе. Задняя кишка с анальным отверстием отсутствуют, поэтому выведение продуктов пищеварения осуществляется в обратном направлении через рот. Строение системы у представителей типа варьирует, что, главным образом, определяется размером животного. При небольших размерах червя кишечник имеет вид слепозамкнутой трубки, при этом всосавшиеся питательные вещества легко диффундируют ко всем участкам тела. У более крупных форм, в
условиях отсутствия кровеносной системы, возникает проблема доставки питательных веществ к удаленным от кишечника органам и тканям, поэтому средняя кишка образует ответвления во все стороны тела, которые доставляют питательные вещества к органам и тканям животного.
Усвоение пищи в значительной мере осуществляется за счет внутриклеточного пищеварения,
осуществляемого эпителиальными клетками средней кишки. Некоторые примитивные формы,
например, бескишечные турбеллярии не имеют кишки, поэтому усвоение пищи у них осуществляется специализированными пищеварительными клетками-фагоцитами,
расположенными в толще паренхимы. Возможно и наружное пищеварение. Так, например, у
турбеллярий в случае если добыча имеет жесткий панцирь, из глотки червя наружу выделяются пищеварительные ферменты, расщепляющие ткани жертвы вне организма, после чего размягченная пища всасывается глоткой. У паразитических форм пищеварительная система редуцирована, и питательные вещества всасываются через поверхность тела.
Утурбеллярий рот располагается на брюшной стороне тела. Бескишечные турбеллярии не имеют средней кишки, глотка может быть развита или отсутствовать. У прочих ресничных червей глотка хорошо развита, ее форма может быть простая, складчатая или бульбовидная.
Утрематод ротовое отверстие располагается на переднем конце тела и окружено ротовой присоской. Глотка окружена мышцами, которые участвуют во втягивании и проталкивании пищи.
Далее следует пищевод и средняя кишка, которая, как правило,бифурцирует (рис. 5).
Рис. 5. Пищеварительная система сосальщиков:
1 – рот, 2 – ротовая присоска, 3 – глотка, 4 – пищевод, 5 – брюшная присоска, 6 – 2 слепо заканчивающиеся кишки