

Филогенез нервной системы
.pdfТЕМАТИЧЕСКИЙ БЛОК №17
Тема: Филогенез систем органов животных и человека. Нервная система.
Цель: Знать основные этапы развития нервной системы у беспозвоночных животных.
Уметь охарактеризовать направления эволюции головного мозга позвоночных. Получить представление о филонтогенетических пороках развития нервной системы у человека.
Основные вопросы темы:
1.Особенности строения нервной системы беспозвоночных животных.
2.Особенности строения ихтиопсидного, зауропсидного и маммального типа мозга.
3.Характеристика строения и функций головного мозга надкласса рыб, классов амфибий,
рептилий, птиц и млекопитающих.
4.Особенности строения головного мозга человека, явление функциональной асимметрии головного мозга у человека.
5.Понятие о филонтогенетических пороках развития нервной системы у человека.
Содержание темы
Нервная система — это совокупность специальных структур, объединяющая и
координирующая деятельность всех органов и систем организма в постоянном
взаимодействии с внешней средой.
Функции нервной системы:
Восприятие различных сигналов (раздражителей) внешней и внутренней среды организма.
Анализаторная – анализ поступающей по сенсорным системам информации.
Моторная – организация ответных движений.
Интегративная – объединение, связь различных сенсорных и моторных структур нервной системы, формирование временных связей, обучение, организация психических процессов.
Принимая прямое участие в формировании ответных реакций на изменения в среде, нервная система выполняет функции регуляции гомеостаза. Кроме того,
благодаря нервной системе осуществляется адекватное взаимодействие организма с окружающей средой не только через организацию ответных реакций эффекторными системами, но и через ее собственные психические реакции — эмоции, мотивации,
сознание, мышление, память, высшие познавательные и творческие процессы.

Нервная система делится на центральную и периферическую. Центральная нервная система образована головным и спинным мозгом. Периферическая нервная система представлена 12 парами черепных и 31 парой спинномозговых нервов с их корешками, ветвями и нервными окончаниями, а также нервными узлами или ганглиями. Часть периферической нервной системы, иннервирующая скелетную мускулатуру, называется соматической нервной системой. Другая часть периферической нервной системы, отвечающая за иннервацию внутренних органов,
кровеносной и эндокринной систем, регуляцию обменных процессов, называется
вегетативной, или автономной нервной системой. Вегетативная нервная система делится на парасимпатическую и симпатическую.
Структурно-функциональной единицей нервной системы является нервная клетка – нейрон. Его основными свойствами являются возбудимость и проводимость.
Нейроны состоят из тела и отростков — коротких, ветвящихся (дендритов) и длинного
(аксона). Аксоны покрыты леммоцитами (шванновскими клетками1), образующими защитную миелиновую оболочку2, которая служит для электрической изоляции и позволяет ускорять передачу нервных импульсов. Между шванновскими клетками находятся узлы Ранвье. Импульсы всегда движутся по дендритам к клетке, а по аксону
— от клетки (рис. 1).
Рис. 1. Строение нейрона:
1 – тело нейрона, 2 – ядро, 3 – дендриты, 4 – Шванновская клетка, 5 – миелиновая оболочка, 6 – узлы Ранвье, 7 – аксон, 8 – ветви аксона
В зависимости от числа и расположения отростков нейроны делятся на униполярные, биполярные и мультиполярные (рис. 2).
1Описаны немецким физиологом Теодором Шванном в 1838 году и названы в его честь.
2Миелиновая оболочка длинных нервов была открыта и названа немецким патологоанатомом Рудольфом Вирховым в 1854 году. Французский патолог и анатом Ранвье позже обнаружил перехваты, или разрывы, в миелиновой оболочке, которые были названы его именем.

Рис. 2. Типы нейронов:
а – мультиполярный, б – биполярный, в – униполярный Моторные, или эфферентные, нейроны передают импульсы от ЦНС к
эффекторам, например, к мышцам. Нейроны, передающие импульсы в центральную нервную систему (ЦНС), называются сенсорными или афферентными. Те и другие нейроны могут связываться между собой с помощью вставочных нейронов
(интернейронов) (рис. 3).
Рис. 3. Функциональные классы нейронов:
а – эфферентный, б – интернейрон, в - афферентный Нейроны связаны между собой синапсами – пространством между соседними
клетками, в котором осуществляется химическая передача нервного импульса с одного нейрона на другой. Синапсы могут возникать между аксоном одного нейрона и телом другого, между аксонами и дендритами соседних нейронов, между одноименными отростками нейронов.
Импульсы в синапсах передаются с помощью биологически активных веществ – нейромедиаторов. Роль медиаторов выполняют норадреналин, ацетилхолин, серотонин,
дофамин и др. Молекулы медиаторов в результате взаимодействия с клеточной мембраной меняют ее проницаемость для ионов Са2+, К+ и Сl-. Это приводит к

возбуждению нейрона. Распространение возбуждения связано с таким свойством нервной ткани, как проводимость (рис. 4).
Рис. 4. Схема химической передачи нервного импульса в синапсе Нервная регуляция осуществляется рефлекторно. Нервный импульс, возникший
при раздражении, проходит определенный путь, называемый рефлекторной дугой.
Простейшая рефлекторная дуга чаще всего состоит из следующих звеньев: рецептор –
нервное окончание, воспринимающее раздражение (находится в органах, мышцах,
коже и т.д.), чувствительный нейрон, передающий импульс в ЦНС, вставочный нейрон,
лежащий в ЦНС (головном или спинном мозге), исполнительный (двигательный)
нейрон, передающий импульс к исполнительному органу или железе. Соматические рефлекторные дуги осуществляют двигательные рефлексы. Вегетативные рефлекторные дуги координируют работу внутренних органов.
Рефлекторная реакция заключается не только в возбуждении, но и в
торможении, т.е. в задержке или ослаблении возникшего возбуждения. Взаимосвязь возбуждения и торможения обеспечивают согласованную работу организма.
ЭВОЛЮЦИЯ НЕРВНОЙ СИСТЕМЫ БЕСПОЗВОНОЧНЫХ
Одноклеточные организмы не обладают нервной системой, однако способны поддерживать «жизненные» функции организма и осуществлять контроль поведения в конкретной среде. Так, например, бактерии посредством большого количества хеморецепторов «чувствуют» питательные вещества или токсины, а наличие механорецепторов позволяет определять препятствия в своей среде. Другие бактерии обладают светочувствительными пятнами, которые заставляют их двигаться к солнечному свету. Получаемая из окружающей среды информация интегрируется и через цепочку сложных химических реакций управляет их движением. У некоторых бактерий (Escherichia coli) есть мини-память, благодаря которой они могут сравнивать входящую информацию с предыдущей информацией, и это «знание» определяет их последующее поведение. Таким образом, даже у самых примитивных организмов имеются три основных компонента для адаптивного управления поведением:
сенсорный, двигательный, также связь между хранением и обработкой информации.

Одноклеточные эукариоты демонстрируют гораздо более сложный контроль над поведением, чем бактерии. Представители простейших (Protozoa) обладают раздражимостью, т. е. способностью отвечать той или иной реакцией на факторы,
действующие извне. Они реагируют на механические, химические, термические,
световые, электрические и иные раздражения. Реакции простейших на внешние раздражения часто выражаются в изменении направления движения и носят название таксис. Таксисы могут быть положительными, если движение осуществляется в направлении раздражителя, и отрицательными, если оно осуществляется в противоположную сторону. Кроме того, многие простейшие обладают специальными органеллами (жгутики, реснички, псевдоподии), благодаря которым осуществляются скоординированные движения за счет активации потенциалзависимых гиперполяризирующих калиевых и деполяризирующих кальциевых каналов.
Так как у простейших нет пространственной разобщенности между раздражителем и реакцией, клетка функционирует одновременно и как рецептор и как эффектор. Появление многоклеточности привело к пространственной разобщенности между раздражителем и реакцией. Роль посредника между рецепторами и эффекторами взял на себя нейрон – нервная клетка. На самых ранних этапах эволюции тела нейронов располагались на поверхности тела животного, а отростки – аксоны – уходили вглубь.
Дальнейшая эволюция зависела от развития нервных цепей, которые состояли уже из нескольких нейронов.
Многоклеточные животные подразделяют на подцарства Parazoa, Mesozoa,
Eumetazoa (рис. 5).
К Parazoa относят тип губок (Porifera = Spongiа), к Mesozoa тип пластинчатых
(Placozoa). Eumetazoa (настоящие многоклеточные) подразделяются на два раздела:
радиально симметричных, или двухслойных (Radiata, Diploblastica), и билатеральных,
или трехслойных (Bilateria, Triploblastica). Радиально симметричные включают два типа: стрекающие животные (Cnidaria) и гребневики (Ctenophora). Все остальные типы животных относятся к билатериям, которые, в свою очередь, подразделяются на две большие группы первичноротые (Protostomia) и вторичноротые (Deuterostomia).
Рис. 5. Формы многоклеточных организмов
Для беспозвоночных животных характерно наличие нескольких источников происхождения нервных клеток. У одного и того же типа животных нервные клетки могут одновременно и независимо происходить из трех разных зародышевых листков.
Полигенез нервных клеток беспозвоночных является основой разнообразия медиаторных механизмов их нервной системы.
Эволюция нервной системы беспозвоночных идет по пути концентрации нервных элементов, в направлении усложнения структурных взаимоотношений в пределах ганглиев. Как специализированная система интеграции на пути структурного и функционального развития, нервная система беспозвоночных проходит через несколько этапов:
диффузная нервная система (нервная сеть);
лестничная нервная система (ортогон);
ганглионарная нервная система (брюшная нервная цепочка).
Диффузная нервная система (нервная сеть) представляет собой скопление мультиполярных и биполярных нейронов, отростки которых могут перекрещиваться,
прилегать друг к другу и лишены функциональной дифференциации на аксоны и дендриты. Нервная сеть может быть локализована в эктодерме и энтодерме.
Тип Кишечнополостные (Coelenterata, Radiata). Представители типа являются двухслойными животными. Их тело представляет собой полый мешок, внутреннее пространство которого является пищеварительной полостью. Нервная система кишечнополостных принадлежит к диффузному типу. Каждая нервная клетка в ней длинными отростками соединена с несколькими соседними, образуя нервную сеть.
Нервные клетки кишечнополостных не имеют специализированных поляризованных отростков. Их отростки проводят возбуждение в любую сторону и не образуют длинных проводящих путей. Контакты между нервными клетками могут быть нескольких типов. Существуют плазматические контакты, обеспечивающие непрерывность сети. Появляются и щелевидные контакты между отростками нервных клеток, подобные синапсам. Такого рода контакты подразделяют на симметричные синапсы (синаптические пузырьки располагаются по обе стороны контакта) и
несимметричные синапсы (в них везикулы располагаются только с одной стороны щели).
Нервные клетки типичного кишечнополостного животного гидры равномерно распределены по поверхности тела, образуя некоторые скопления в районе ротового отверстия и подошвы (рис. 6). Диффузная нервная сеть проводит возбуждение во всех направлениях. При этом волну распространяющегося возбуждения сопровождает волна мышечного сокращения.

Рис. 6. Схема строения нервной системы гидры Лестничная нервная система (ортогон) представляет собой упорядоченную
структуру, которая состоит из ассоциативных и двигательных клеток, формирующих вместе несколько пар продольных тяжей, или стволов, соединенных большим числом поперечных и кольцевых комиссуральных стволов. Концентрация нервных элементов сопровождается их погружением в глубь тела. Появление ортогонального типа нервной системы является результатом централизации и концентрации нервных элементов.
Представлена у плоских и круглых червей.
Тип Плоские черви (Plathelminthes). Представители этого типа являются трехслойными животными в связи с появлением третьего зародышевого листка– мезодермы. У примитивных плоских червей нервные клетки образуют нервную систему, мало отличающуюся от нервной системы стрекающих кишечнополостных. У
более продвинутых форм в головной части имеется скопление нервных клеток
(церебральные ганглии), что можно считать первым шагом на пути формирования центральной нервной системы. От ганглиев отходят продольные (латеральные)
нервные тяжи, соединенные между собой поперечными нервными волокнами. Хемо- и
механорецепторы распределены по поверхности тела. На головном конце имеется группа вкусовых клеток и небольшие ямки с расположенными в них фоточувствительными клетками (рис. 7).
Рис. 7. Схема строения нервной системы плоских червей (белой планарии)
При общей ограниченности форм поведения плоские черви демонстрируют простейшие ориентировочные рефлексы. У паразитических форм наблюдается частичная или полная редукция органов чувств и нервных клеток.
Тип круглые черви (Nemathelminthes) – трехслойные животные, имеющие первичную полость тела. Их нервная система построена по типу ортогона. Основной группой типа является класс круглых червей (Nematoda). Центральная часть их нервной системы образована окологлоточным нервным кольцом, окружающим переднюю часть пищевода. С кольцом связана пара боковых головных ганглиев. От кольца вперед отходят 6 коротких нервных веточек, а назад направляются 6 нервных стволов. Среди последних два, проходящие по срединной спинной и брюшной линиям в соответствующих валиках гиподермы, наиболее мощные. Оба главных нервных ствола соединяются между собой многочисленными комиссурами, опоясывающими тело в виде полукольца попеременно с правой и левой сторон. Мускульный слой разбит валиками гиподермы на 4 продольные ленты. Спинной нервный ствол иннервирует мышцы обеих боковых спинных лент, брюшной - брюшных. Органы чувств нематод развиты слабо: имеются осязательные бугорки или щетинки, хеморецепторные органы,
а также примитивные глаза - пигментные пятна или слабо развитый бокал.
Ганглионарная нервная система характеризуется тем, что большая часть нервных клеток собраны в нервные узлы — ганглии. Соединенные между собой ганглии, в свою очередь, формируют нервную цепочку, расположенную на брюшной стороне тела. Характерна для аннелид, членистоногих и моллюсков.
Тип Кольчатые черви (Annelida) – трехслойные целомические животные со сквозным кишечником и сегментированным телом. Нервные клетки сгруппированы в ганглии, образующие ЦНС. Ганглии головной части тела образуют мозг. В каждом метамере (сегменте тела) ганглии расположены попарно и билатерально, соединяясь комиссурами через среднюю линию (сходство с ортогоном). Соединение между метамерами осуществляется с помощью продольных тяжей, или коннектив (гигантские нервные волокна, образующиеся при слиянии нескольких аксонов), а с периферией - с
помощью нервов. Ганглии, соединенные коннективами, образуют брюшную нервную цепочку (рис. 8). Подглоточные ганглии контролируют все жизненно важные рефлексы животного. Органы чувств представлены сенсиллами, которые представлены механорецепторами, хеморецепторами и фоторецепторами. Тенденция к концентрации нервных клеток и чувствительных рецепторов на переднем конце тела называется
цефализацией.
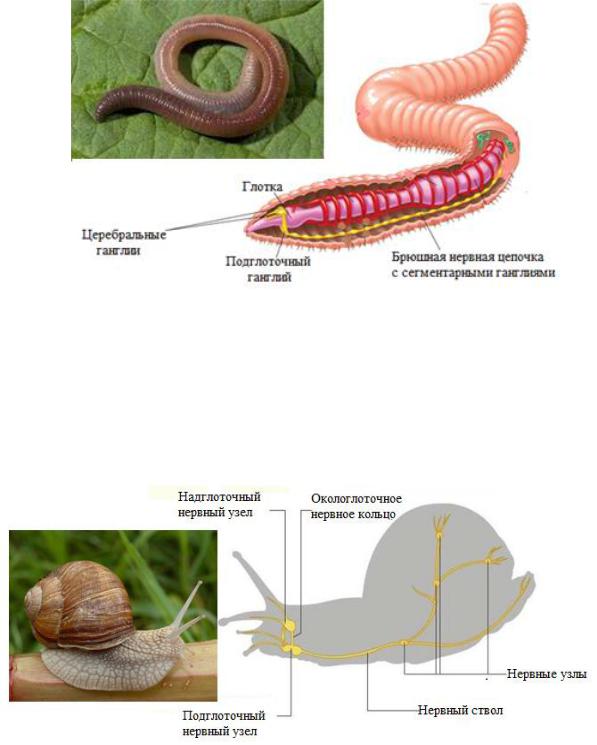
Рис. 8. Схема строения нервной системы кольчатых червей
Тип Моллюски (Mollusca). Нервная система моллюсков может быть различной степени сложности и состоит из ганглиев, наиболее крупные из которых сгруппированы вокруг пищевода. Пары ганглиев соединяются между собой комиссурами, а с другими ганглиями – коннективами (рис. 9).
Рис. 9. Схема строения нервной системы моллюсков (Брюхоногие моллюски)
Из органов чувств имеются статоцист (механорецепторный орган равновесия) и
осфрадий (орган химического чувства), у многих хорошо развиты глаза. Центральная нервная система головоногих моллюсков достигает большой степени развития.
Расположенные вокруг пищевода ганглии сильно увеличены и, сливаясь, образуют настоящий мозг. Очень высокого уровня развития достигают глаза. Зрительные ганглии превращаются в сложные зрительные доли – самые крупные отделы мозга. Нейроны этих долей дифференцируются на ряд форм, отличающихся от униполярных нейронов,
характерных для беспозвоночных.

Тип Членистоногие (Arthropoda). К данному типу относятся животные,
имеющие хитиновый наружный скелет и членистые конечности. Нервная система членистоногих в исходном состоянии устроена так же, как у кольчатых червей. В ходе прогрессивного развития членистоногих строение нервной системы усложняется.
Во-первых, у членистоногих в большинстве случаев нервная лестница сменяется брюшной нервной цепочкой, то есть происходит сближение (или даже слияние)
ганглиев в пределах одного сегмента. В исходном состоянии ганглии брюшной нервной цепочки (лестницы) сохраняют свою приуроченность к "родному" сегменту.
Во-вторых, часто происходит продольная концентрация ганглиев,
проявляющаяся в сближении (слиянии) ганглиев разноименных сегментов. В первую очередь в этот процесс вовлекаются ганглии передних сегментов. Как правило, за счет слияния ганглиев второго, третьего и четвертого сегментов формируется крупный ганглий, часто именуемый подглоточным.
В-третьих, у представителей типа формируется крупный головной мозг (его часто называют надглоточным или супраэзофагальным ганглием), состоящий из трех отделов: протоцеребрум, дейтоцеребрум и тритоцеребрум. В протоцеребруме находятся ассоциативные зоны (центральное тело и грибовидные тела3) и зрительные центры. В дейтоцеребруме располагаются антеннальные центры и «обонятельные» доли. В тритоцеребруме находятся центры, регулирующие работу ротового аппарата.
Подглоточный ганглий служит главным моторным центром и необходим для координированной ходьбы, плавания или полета (рис. 10).
Рис. 10. Общая схема строения нервной системы членистоногих
Периферическая нервная система членистоногих представлена нервами,
отходящими от головного мозга и от ганглиев брюшной нервной цепочки. В исходном
3Грибовидные тела, или corpora pedunculata являются высшим «интеллектуальным» центром беспозвоночных. Организованы по принципам, общим с корой головного мозга млекопитающих. В коре и грибовидных телах нервные клетки расположены слоями или стратифицированы.