

17.6. Адаптация в слуховой системе.
Явление адаптации присуще всем сенсорным системам, и слуховая система не составляет исключения. Это свойство как периферических рецепторов, так и центральных нейронов. Адаптация к данному уровню звука приводит к повышению слухового порога, но из этого не следует, что она нежелательна или бесполезна. Адаптация к некоторым интенсивностям и частотам может в действительности понизить дифференциальный порог, усилив различение разных уровней слухового давления . Так, например, тон в 4000 Гц на рис 5-6 при 33 дБ УЗД имеет уровень громкости 40 фонов, а тот же тон при 52 дБ УЗД-60 фонов. Следовательно, в этой части графика, чтобы получить разницу уровня громкости в 20 фонов, нужно усиление звукового давления на 19 дБ. Но если ухо адаптировано к долгому воздействию 4000 Гц, то изофоны в этой области смещаются вверх; в частности, для низких уровней громкости изофоны сместятся дальше, чем для высоких . В результате разница в уровне громкости в 20 фонов потребует изменения меньше чем на 19 дБ УЗД, например на 14 дБ. Иными словами, в адаптированном состоянии данная разница в уровне звукового давления вызовет большую разницу в уровне громкости. Субъективная разница становится больше, и, следовательно, порог различения понижается.
Предыдущее изложение могло создать впечатление, что ощущение тона основано полностью на локализации смещения в улитке, зависимой от частоты, как это постулирует теория места, или на настройке волокон слухового нерва на их характеристические частоты. Однако не все психофизические наблюдения над тональными ощущениями можно объяснить одной только теорией места. Так, например, при стимуляции уха последовательностью коротких импульсов давления (щелчков) с интервалами в 5 мс, как и следовало ожидать, слышится тон в 200 Гц. Но если предварительно профильтровать звук так, чтобы пропускались частоты только выше определенного уровня (скажем, 600 Гц), то в стимуле уже не будет частоты 200 Гц. Однако субъективно воспринимаемый тон по-прежнему будет 200 Гц. Это явление еще не получило убедительного объяснения, но его нельзя объяснить одной только теорией места. По-видимому, мозг "воссоздает" недостающую основную частоту информации, которая содержится в сохранившихся обертонах.
Лекция 18. Физиология чувства равновесия
18.1. Анатомия и физиология периферического органа. Рецепторы органа равновесия и стимулы, их возбуждающие.
В височной кости близ улитки лежит вестибулярный орган-преобразователь, тесно связанный с поддержанием равновесия. Относительная ориентация этих частей лабиринта была показана на рис.17.2.. Вестибулярный орган филогенетически связан с ухом, в особенности в том, что касается рецепторов обеих систем. В том и другом случае рецепторами являются волосковые клетки. Адекватным стимулом для волосковых клеток служит сгибание цилий. То же самое относится к вестибулярному органу, но здесь смещающее усилие возникает иначе. На рис.18.1. показано действие этого механизма в одном из сенсорных образований вестибулярного органа -макуле.
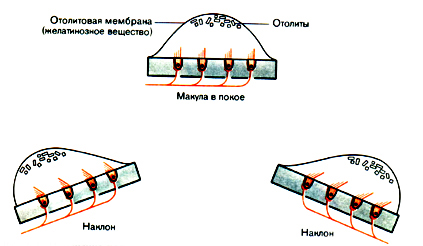
Рис.18.1. Изменение положения макулярного органа при наклоне головы (адекватный для этого органа стимул) в двух направлениях.
Цилии волосковых клеток вдаются в желатинозную массу, в которую погружены мелкие гранулы с высоким удельным весом. Масса вместе с ее содержимым называется отолитовой мембраной. Цилии разной длины образуют лесенку, причем самая длинная из них (называемая киноцилией) построена иначе, чем остальные. Такая дифференциация отличает эти волосковые клетки от клеток в улитке, которые лишены ки-ноцилии и обладают только стереоцилиями.
В верхней части рис. видна макула в покое. При наклоне головы в ту или другую сторону сила тяжести слегка смещает отолитовую мембрану, как показано на двух нижних рисунках. Смещаясь на это небольшое расстояние относительно сенсорного эпителия, она сгибает цилии. Сгибание служит стимулом для рецепторов. Афферентный нерв передает сигнал об этом состоянии в ЦНС следующим образом. Волокна активны (активность покоя) даже в нейтральном положении. При сгибании цилий в одном направлении частота разряда возрастает, при сгибании в другом - снижается. Усиление активности афферентного нервного волокна происходит, когда смещающее усилие в пучке цилий направлено в сторону киноцилий; торможение наступает при смещении в противоположную сторону. Таким образом, главным эффективным стимулом макулы служит сила тяжести. Она соответствует специальному случаю линейного ускорения. Макулы отвечают также на все другие формы линейного ускорения, например такого, какое возникает при пуске в ход или при торможении автомобиля. В таких случаях отолитовая мембрана смещается по отношению к лежащему под ней сенсорному эпителию точно так же, как подвижный предмет скользит вперед при внезапном включении тормозов автомобиля.