

Лекция 4. Распространение электрона и проведение потенциала действия
Мы уже говорили о том, что если мембрана нервного волокна (нервной клетки или мышечного волокна) в какой-то точке подвергается действию входящего или выходящего постоянного электрического тока, то в этой точке МП оказывается постоянно измененным - сниженным при выходящем токе и повышенным при входящем токе. Это состояние измененного МП называют физическим электротоном (тонус, т. е. постоянное напряжение).
Физический электротон даже при строго локальной подаче тока с помощью внутриклеточного микроэлектрода на какую-либо точку мембраны не локализуется в этой точке мембраны, расползается во все стороны от нее, ослабевая (декрементируя). Это распространение физического электротона по мембране, хотя само по себе и не обеспечивает дальней связи, тем не менее имеет очень большое физиологическое значение. Оно обеспечивает ближнюю связь по мембране в объектах с малой протяженностью (например, в телах нейронов, в некоторых мышечных волокнах, в клетках глии, эпителия и др.). Это же распространение электротона является одной из основ механизма бездекрементного проведения возбуждения (потенциала действия) в протяженных нервных и мышечных волокнах.
Рассмотрим некоторые подробности распространения электротона и причины этого распространения.
При включении, достаточно длительном действии, а затем выключении тока электротон соответственно возникает (нарастает, расползается), далее существует как стационарное распределение потенциалов вокруг электрода и, наконец, исчезает. Причина распространения электротона заключена в кабельной структуре клеточных объектов, например нервного (мышечного) волокна. Нервное волокно подобно кабелю имеет хорошо проводящий электрический ток стержень - аксоплазму, и плохо проводящую высокоомную оболочку - мембрану. При такой структуре электрический ток, вводимый микроэлектродом в какую-либо точку кабеля (при наружном расположении другого электрода) как бы стремится использовать проводящий стержень для подхода к возможно большему числу точек высокоомной мембраны, сопротивление которой ему необходимо преодолеть. Очевидно, если сопротивление аксоплазмы (Ri) и сопротивление среды (Re) по сравнению с сопротивлением мембраны (Rм) пренебрежимо малы, то подаваемый ток сможет использовать все точки мембраны волокна практически равномерно и элек-тротон равномерно охватит все волокно. Наоборот, при сопротивлении мембраны (RM) пренебрежимо малом по сравнению с сопротивлением аксоплазмы (Ri) и среды (Re), электротон локализовался бы практически в области микроэлектрода. При соизмеримости RM и Re, стационарный электротон экспоненциально затухает в пространстве с константой
![]()
где RM, Ri и Re рассчитаны на единицу длины (1 см).
Так как Ri зависит от диаметра волокна (D), то при использовании тех же показателей, рассчитанных на единицу площади (1 см2), имеем:
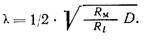
Постепенное, экспоненциальное развитие и исчезновение электротона во времени (в каждой точке мембраны) объясняется наличием мембранной емкости См. Скорость этого развития определяется постоянной времени мембраны τм. Чем больше τ, тем медленнее возникает и исчезает электроток. Вдали от точки приложения электрода электроток возникает и исчезает медленнее. Поскольку электроток при включении тока как бы расползается по мембране, можно говорить о скорости распространения электротона Vэт.
Обратимся теперь к феномену проведения возбуждения, т. е. распространения ПД, имеющему место в нервном или фазном мышечном волокне, а также в соме и дендритах нейрона.
Возбуждение (ПД) как и электротон, распространяется по мембране клеток всесторонне (в волокнах, грубо говоря, "двухсторонне"). Но проведение возбуждения в отличие от распространения электротона происходит (в однородном проводнике!) без снижения амплитуды ПД и без снижения скорости (бездекрементно). Механизм проведения возбуждения, как хорошо теперь известно, включает в себя два компонента: 1) раздражающее действие катэлектротонического сигнала, порождаемого локальным ПД, на соседний участок электровозбудимой мембраны и 2) возникновение ПД в этом соседнем раздражаемом участке мембраны. Проведение ПД - это нечто вроде эстафеты, в которой каждый участок вдоль волокна выступает как раздражаемый, а затем как раздражающий последующий участок.
То, что ПД распространяется с помощью петли электротона, доказывается фактом перескока нервного импульса через узкий участок инактивированной мембраны (отравленной или охлажденной) . Хотя сам по себе этот участок не генерирует ПД, но ПД, возникающий в доблоковой области, вызывает за участком блока катэлектротонический сдвиг потенциала, на вершине которого можно определить ЛО. Иногда здесь получается и ПД В последнем случае возбуждение как бы "перескакивает" через инактивированный участок и распространяется дальше. Перескоку возбуждения через такой участок отчетливо способствует снижение сопротивления внешней среды (Re), в частности, шунтировка нервного кабеля с помощью металлических стерженьков и ртути, применявшаяся Ходжкиным в опытах на гигантском аксоне кальмара (1939). Снижение Re делает электроток более распространенным в пространстве что и способствует перескоку ПД.
Назовем реальные скорости в некоторых немиелинизированных нервных проводниках. В гигантском аксоне кальмара V = 25 м/с (при D= 0,5 мм). В тонких волокнах позвоночных - около 1 м/с (при D=l мкм).
Остановимся на понятии "протяженности возбужденного участка", т. е. участка, в котором в данный момент генерируется пик ПД (та или иная его стадия) . Протяженность возбужденного участка L=VПД • tПД. В гигантском аксоне кальмара L=l мс • 25 мм/мс = 25 мм. В тонких немиелинизированных волокнах при tПД= 1 мс: V = 1 мм/мс; L=l мс •1 мм/мс=1 мм. Проведение возбуждения в миелинизированных нервных волокнах, характерных для скоростных линий нервной связи у высших животных, осуществляется тем же способом, что и в немиелинизированных, но здесь имеются некоторые существенные особенности. Миелиновые муфты, каждая из которых формируется намоткой одной шванновской клетки на аксон, являются хорошими электроизоляторами: у миелина (R = 0,16 МОм•см2) С= 0,005 мкФ/см2. Поэтому в миелинизированном волокне проводят электрические токи и генерируют ПД в ответ на них по существу только узкие (∼ 1 мкм) оголенные участки, расположенные между муфтами - перехваты Ранвье, что было показано Тасаки и Такеути в 1942 г. и Хаксли, Штемпфли в 1949 г. Распространение ПД здесь осуществляется скачкообразно - "сальтаторно" - от перехвата к перехвату. Поскольку миелиновые сегменты гораздо протяженнее перехватов (1000 - 2000 мкм против 1 мкм), то такая форма функционирования проводника экономична в смысле расхода ионов и нагрузки на ионный насос и обеспечивает существенно большие скорости проведения возбуждения (рис.4.1. ).
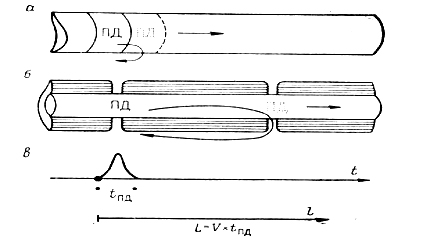
Рис. 4.1. Сопоставление непрерывного и сальтаторного распространения потенциала действия.
Схемы: а - непрерывного распространения ПД в немиелинизированном волокне ;
б - сальтаторного проведения в миелинизированном нервном волокне;
в - развитие одиночного пика ПД во времени и распространение его в пространстве.
VПД здесь иначе зависит от диаметра волокна, чем в случае немиелинизированных проводников. Эмпирически установлено, что в миелинизированных волокнах позвоночных VПД=КD; ν линейно зависит от D, потому что ν пропорциональна длине межперехватного участка l, а l линейно зависит от D (при этом время на раздражение очередного перехвата оказывается приблизительно постоянным). "К" варьирует от 6 до 1,73 для разных групп волокон. У млекопитающих в группе наиболее толстых миелинизированных A-волокон К=6, т. е у волокон с D= 20 мкм VПД=120 м/с (при этом L = 60 MM), a у волокон с D=4 мкм VПД=24 м/с (L - 24 мм). Обратим внимание на то, что в последнем случае скорость проведения приблизительно соответствует достигаемой в гигантском аксоне кальмара (D = 500 мкм). Это наглядно демонстрирует прогрессивность миелинизации, позволяющей достигать высоких скоростей нервнойсигнализации при малой толщине нервных проводников. И понятно, что нервные болезни, при которых происходит демиелинизация, характеризуются сильными расстройствами функций нервной системы.
В заключение отметим особенности проведения ПД в участках резкого изменения свойств нервного проводника. Таковым является, например, переход миелинизированного нервного волокна в оголенное и разветвленное окончание. Площадь мембраны довольно протяженного окончания обычно значительно (на 2 порядка) больше площади перехвата Ранвье (хотя окончание тоньше магистрального волокна). При таком соотношении площадей проведение ПД из перехвата в окончание (например, моторное) несколько замедленно и имеет сниженный ГФ. Это объясняется падением плотности локального раздражающего тока в окончании. Проведение же из окончания (например, сенсорного) в перехват ускорено и облегчено из-за концентрации тока ПД окончания в перехвате.
Аналогичная ситуация наблюдается в участках разветвления магистральных нервных волокон (а также проводящих ПД дендритов).
Поскольку рабочая площадь мембраны в неразветвленной части меньше, чем в разветвленной, то проведение ПД из первой во вторую происходит с некоторым замедлением и при сниженном ГФ]. Наоборот, синхронные ПД ветвей взаимно облегчают себе вход в неразветвленную часть проводника. Наконец, сходные явления имеют место в участках резкого изменения диаметра проводника, например, в районе перехода тела нейрона в начальный сегмент аксона или дендрит. Здесь взаимодействующие соседние участки мембраны также имеют существенно разную по величине рабочую площадь. И это обстоятельство при прочих равных условиях способствует проведению ПД из расширенного сегмента в суженный (например, из сомы в дендрит) и затрудняет проведение в противоположном направлении.