

2 курс / Нормальная физиология / Физиология_человека_Семенович_А_А_,_Переверзев_В_А_
.pdfРис. 10.6. Последовательность включения ферментных комплексов в тканевое дыхание
мембран убихинон (кофермент Q) и группа гемсодержащих белков (цитохромов b, c, a, a3). В системе переноса электронов важна роль железа, которое включается в состав ферментов в структуре гема (в цитохромах) или в составе комплекса FeS.
Завершаюшим этапом работы дыхательной цепи является реакция, катализируемая ферментом цитохромоксидазой, ко! торая через свой кофермент а3 передает электроны непосред! ственно кислороду и последний взаимодействует с протонами с образованием воды. Молекула кислорода принимает четыре электрона и формирует две молекулы воды.
Во время переноса электронов комплексы дыхательной цепи (1, 3 и 4!й) перекачивают протоны из матрикса в межмембранное пространство и на внутренней мембране возникает градиент протонов (градиент электрохи! мического потенциала). Этот градиент используется особым комплексом ферментов (называемым АТФ!синтетазой) для синтеза АТФ. Для синте! за и переноса одной молекулы АТФ из митохондрии в цитоплазму исполь! зуется энергия движения четырех протонов по электрохимическому гра! диенту через внутреннюю мембрану митохондрии. Поскольку в процессе образования двух молекул воды в межмембранное пространство митохон! дрии переносится 20 протонов, то их энергии хватает на синтез пяти моле!
361
кул АТФ (20:4=5). Возможно функционирование и укороченного варианта дыхательной цепи, когда переносится только 12 протонов и синтезируется только три молекулы АТФ.
Такой механизм синтеза АТФ за счет энергии градиента электрохимического потенциала получил название окисли! тельного фосфорилирования и составляет основу получения АТФ в аэробных условиях. Образованная таким путем АТФ является основным источником энергии для протекания жиз! ненных процессов у высокоорганизованных живых существ.
Сопряжение между переносом электронов и синтезом АТФ может нарушаться в присутствии некоторых химических соединений или при возникновении условий, повышающих проницаемость внутренней мем! браны митохондрий для протонов. В этом случае протоны переходят в матрикс, минуя АТФ!синтетазу, синтез АТФ замедляется. Энергия пере! носа элетронов высвобождается в форме тепла, а клетки испытывают энергетический голод. Такие события получили название разобщение окислительного фосфорилирования, а вещества, которые его вызы! вают, – разобщители. Например, сильнодействующим разобщителем является 2, 4!динитрофенол. В митохондриях бурой жировой ткани роль разобщителя выполняет специальный белок – термогенин. Митохонд! рии, содержащие термогенин, являются хорошими источниками тепла и помогают приспосабливаться к низким температурам.
Известно большое количество ингибиторов переноса элек! тронов по дыхательной цепи. Они являются сильнодействую! щими ядами (цианистый водород и его производные). Их дей! ствие вызывает прекращение переноса электронов и, следова! тельно, прекращение дыхания и смерть.
Молекула кислорода может присоединять от одного до четырех элект! ронов. В зависимости от количества принятых электронов возникают раз! ные производные кислорода. Присоединение четырех электронов к моле! куле кислорода завершается образованием воды. Присоединение иного числа электронов к молекуле кислорода приводит в образованию так назы! ваемых активных форм кислорода: супероксидного анион!радикала (один электрон), пероксидного радикала (два электрона) и гидроксильного ради! кала (три электрона). Эти формы кислорода обладают высокой реакцион! ной способностью, и образование их в значительных количествах может оказывать повреждающий эффект на клетку. Это свойство кислорода, в частности, используют макрофаги, генерируя активные формы кислорода для разрушения фагоцитируемых ими микроорганизмов.
362
В пероксисомах оксидоредуктазы также переносят водоро! ды на кислород, однако при этом реакция ведет к образованию пероксида водорода (пероксидного радикала кислорода):
H2 + O2 → H2O2.
Образуются активные формы кислорода и в мембранах эн! доплазматической сети. Обычно количество формирующихся активных форм кислорода находится под контролем специаль! ных антиоксидантных систем. Различают ферментную и не! ферментную антиоксидантные системы. К ферментам, разру! шающим активные формы кислорода, относятся супероксид! дисмутаза, каталаза, глутатионпероксидаза и др. Нефермента! тивно ограничивают действие радиклов кислорода витамины Е, С и А, мочевая кислота и другие соединения.
Еще одной формой потребления кислорода клетками явля! ются процессы гидроксилирования, при которых кислород присоединяется к молекуле, формируя в ней гидроксильную группу. Эти реакции широко используются для борьбы с гид! рофобными молекулами, оказывающими неблагоприятное воздействие (ксенобиотики) на клетки. Их гидрокислирование позволяет в последующем присоединить гидрофильные моле! кулы (глюкуроновую кислоту, сульфат) и, повысив раствори! мость, вывести их из организма почками.
10.7. Регуляция дыхания
Регуляцией дыхания называют процесс управления венти! ляцией легких, направленный на поддержание дыхательных констант внутренней среды организма и приспособление дыха!
ния к изменяющимся условиям внешней и внутренней среды.
Функциональная система регуляции дыхания. Механиз! мы регуляции дыхания объединяются в функциональную сис! тему, деятельность которой направлена на поддержание дыха! тельных констант внутренней среды организма (ее упрощен! ная схема представлена на рис. 10.7). Этими константами (го! меостатическими показателями) являются напряжение кислорода, напряжение углекислого газа и показатель рН кро! ви и ликвора. Таким образом, система регулирует сразу три показателя. Такие системы относят к весьма сложным. Рас! смотрим сначала общую структуру и свойства системы. За ве! личиной регулируемых параметров следят хеморецепторы.
363
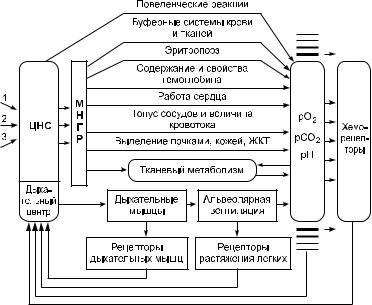
Рис. 10.7. Схема функциональной системы, регулирующей оптимальный |
уровень дыхательных констант внутренней среды организма: |
1–3 – импульсация от экстеро!, интеро! и проприорецепторов |
Импульсы от них идут по каналу обратной связи в центральную нервную систему, прежде всего в дыхательный центр. Этот центр может воспринимать непосредственное воздействие га! зов, переносимых с кровью. К дыхательному центру приходят также импульсы от рецепторов, контролирующих растяжение легких и состояние дыхательных мышц. Таким образом по не! скольким каналам обратной связи жизненно важный отдел ды! хательного центра получает информацию как о величине регу! лируемых показателей, так и о состоянии исполнительных ме! ханизмов, обеспечивающих газообмен. На основе анализа поступающей информации и связей с другими отделами цент! ральной нервной системы формируется комплекс импульсов, передающихся к дыхательным мышцам и многим внутренним органам, которые выполняют роль эффекторов (исполнитель! ных звеньев) в процессах регуляции оптимального уровня ды! хательных констант внутренней среды организма. Среди эф! фекторных структур и механизмов функциональной системы
364
регуляции дыхательных констант выделяют: работу дыхатель! ных мышц, обеспечивающих внешнее дыхание, работу сердца, изменение тонуса сосудов, объема циркулирующей крови, ко! личества эритроцитов и содержания в них гемоглобина, изме! нение выделения кислых или щелочных продуктов почками и желудочно!кишечным трактом, изменение интенсивности и характера метаболизма в тканях.
Таким образом, рассматриваемая функциональная система имеет замкнутый контур регулирования, со многими (не менее четырех) контурами обратной связи и является системой взаи! мосвязанного регулирования трех показателей. Регуляция в ней ведется по отклонению и по возмущению, система способ! на к самообучению. Регуляцию по отклонению можно просле! дить по результатам, полученным исследователями в экспери! ментах на животных: при введении кислот в кровь или ликвор наблюдается гипервентиляция, организм стремится компен! сировать подкисление крови за счет усиления выведения угле! кислого газа через легкие. Регуляция по возмущению ярко видна по резкому увеличению интенсивности внешнего дыха! ния в первые же секунды начала выполнения мышечной рабо! ты. В это время еще нет изменения уровня рО2, рСО2 и рН крови и ликвора, а вентиляция легких резко увеличивается. Организм оценивает приток импульсаций от скелетных мышц как сигнал о возмущающем воздействии, предвещающем сдвиг дыхательных констант, и изменяет работу эффекторных структур так, чтобы предотвратить этот сдвиг. О наличии са! мообучения системы регуляции дыхания свидетельствует воз! можность выработки условных рефлексов, изменяющих ин! тенсивность вентиляции, и ряд других специальных опытов (например, опыты с повторным дыханием через противогаз, когда при повторных подключениях этого устройства дыха! тельный центр все более точно изменяет объем вдоха и выдоха для компенсации подсоединяемого дополнительного мертвого пространства).
В рассматриваемой нами функциональной системе имеют! ся многие блоки, в нее входит ряд рефлекторных реакций и гу! моральных механизмов регуляции функций. Рассмотрим важ!
нейшие из этих составляющих.
Дыхательный центр. Дыхательным центром называют совокупность нейронных структур, расположенных в различ! ных отделах центральной нервной системы, регулирующих
365
ритмические координированные сокращения дыхательных мышц и приспособление дыхания к изменяющимся условиям среды и потребностям организма. Среди этих структур выде! ляют жизненно важные отделы дыхательного центра, без которых дыхательные движения прекращаются. К ним отно! сятся отделы, расположенные в продолговатом и спинном мозге. В спинном мозге в структуру дыхательного центра вхо! дят мотонейроны диафрагмального нерва (3–5!й шейные сег! менты) и мотонейроны межреберных нервов (2–12!й грудные сегменты).
Особое значение имеет отдел дыхательного центра в про! долговатом мозге. Он расположен в области дна 4!го желудоч! ка и представляет собой парное образование, имеющее инспи! раторный (посылает импульсы к мышцам вдоха) и экспиратор! ный (обеспечивает выдох) отделы. В каждом из этих отделов находятся группы нейронов, взаимодействие которых обеспе! чивает формирование частоты и глубины дыхательных движе! ний. В инспираторном отделе имеются нейроны, обладающие автоматией.
Важную роль в определении характера дыхательных дви! жений играют импульсы, приходящие по афферентным волок! нам от рецепторов, а также от коры большого мозга, лимби! ческой системы и особенно гипоталамуса. Схема нервных свя! зей дыхательного центра представлена на рис. 10.8. Рассмот! рим циркуляцию импульсаций в этой системе на протяжении одного дыхательного цикла.
Вначале благодаря автоматии инспираторных нейронов и под влиянием приходящей к ним импульсации от рецепторов, чувствительных к рО2 , рСО2 и рН, а также от других интеро! и экстерорецепторов возбуждаются нейроны инспираторного отдела продолговатого мозга. Импульсы от них идут по нисхо! дящим путям и переключаются на мотонейроны спинного моз! га, формирующие диафрагмальные и межреберные нервы. По этим нервам импульсы приходят к мышцам, обеспечивающим расширение грудной клетки, вслед за которой расширяются легкие и происходит вдох. При вдохе активируются рецепторы растяжения дыхательных путей и легких. Импульсация от ре! цепторов растяжения идет по афферентным волокнам в стволе блуждающего нерва в продолговатый мозг и активирует экс! пираторные нейроны. Так замыкается один контур механизма регуляции дыхания. Второй регуляторный контур также начи!
366

Рис. 10.8. Схема нервных связей дыхательного центра:
Ио – инспираторный отдел дыхательного центра продолговатого мозга; Эо – экс! пираторный отдел, ПТо – пневмотаксический отдел моста; 1 – диафрагмальный нерв; 2 – межреберные нервы; 3 – рецепторы дуги аорты; 4 – рецепторы каро! тидного тельца; 5, 7 – пути передачи импульсов между Ио, Эо и ПТо; 6 – влия! ние высших отделов ЦНС на жизненно важные отделы дыхательного центра; 8, 9 – переключение импульсов от Ио на мотонейроны в шейных и грудных сегментах спинного мозга; 10 – начало афферентных волокон, идущих к Эо в стволе
n. vagus; 11, 12 – эфферентные пути к экспираторным мышцам
нается от инспираторных нейронов и проводит импульсы к нейронам так называемого пневмотаксического отдела дыха! тельного центра, расположенного в мосту мозга. Этот отдел координирует взаимодействие инспираторных и экспиратор! ных нейронов продолговатого мозга. Пневмотаксический от! дел перерабатывает пришедшую от инспираторного центра информацию и посылает поток импульсов, возбуждающих
367
нейроны экспираторного центра. Когда потоки импульсов, приходящих от пневмотаксического отдела и от афферентов вагуса, сходятся на экспираторных нейронах, последние быст! ро возбуждаются и тормозят активность инспираторных ней! ронов по принципу реципрокного торможения. Импульсация к мышцам вдоха прекращается и они расслабляются. Этого до! статочно, чтобы произошел спокойный выдох. При усиленном выдохе от экспираторных нейронов посылаются импульсы, вызывающие сокращение внутренних межреберных мышц и мышц брюшного пресса.
Вышележащие отделы головного мозга оказывают корри! гирующие влияния на нейроны отделов дыхательного центра в продолговатом мозге и мосту. О наличии влияния коры боль! шого мозга на отделы дыхательного центра в спинном и про! долговатом мозге свидетельствует возможность произвольно! го изменения частоты, глубины и задержки дыхания челове! ком. Корковые влияния передаются на эти центры, как через пирамидные, так и экстрапирамидные пути. Кора большого мозга обеспечивает включение дыхательной системы в пове! денческие реакции, речевую функцию, пение.
Гипоталамус играет ведущую роль в изменениях дыхания, свя! занных с реакциями на болевые раздражения, физическую на! грузку, эмоциональное возбуждение, а также обеспечивает во!
влечение дыхательной системы в терморегуляторные реакции.
Рецепторы к кислороду, углекислому газу и показателю рН. Рецепторы, чувствительные к изменениям напряжения кислорода, углекислого газа и водородных ионов, обеспечива! ют непрерывный контроль за уровнем этих гомеостатических констант в артериальной крови и ликворе. Эти рецепторы ак! тивны уже при нормальном уровне рО2, рСО2 и показателя рН и от них идет непрерывная (тоническая) импульсация, спо! собствующая активации инспираторных нейронов.
Рецепторы к кислороду сосредоточены в каротидном тель! це (область бифуркации общей сонной артерии). У животных эти рецепторы имеются и в тельце дуги аорты. На снижение рО2 в артериальной крови клетки каротидного тельца реагиру! ют усилением выделения медиатора допамина. Допамин акти! вирует рецепторные окончания афферентных волокон языко! глоточного нерва. Таким образом, активность рецепторов к кислороду возрастает при снижении его напряжения в артери! альной крови. Импульсы от них возбуждают инспираторные
368
нейроны, и вентиляция легких увеличивается главным обра! зом за счет учащения дыхания.
Рецепторы, чувствительные к углекислому газу, имеются в каротидном тельце и в дуге аорты, а также непосредственно в продолговатом мозге (центральные хеморецепторы, сосредо! точенные в тельцах диаметром около 2 мм) в области выхода подъязычного нерва. Рецепторы к углекислому газу восприни! мают также изменения концентрации ионов Н+. Рецепторы артериальных сосудов контролируют рСО2 и рН плазмы кро! ви. Они активируются при увеличении рСО2, (или снижении рН плазмы). Исходящая от них импульсация вызывает увели! чение вентиляции легких главным образом за счет углубления дыхания. Центральные хеморецепторы контролируют рСО2 и рН ликвора и межклеточной жидкости продолговатого мозга. Они активируются при накоплении углекислого газа или под! кислении ликвора и возбуждают инспираторные нейроны, вы! зывая усиление дыхания.
Чрезмерное увеличение активности рецепторов по рСО2 и рН приводит к возникновению субъективно тягостных, мучи! тельных ощущений удушья, нехватки воздуха. В этом легко убедиться, если сделать длительную задержку дыхания. Важно помнить о том, что вызванная недостатком кислорода в арте! риальной крови стимуляция артериальных рецепторов к кис! лороду не сопровождается субъективно неприятными ощуще! ниями. В условиях, когда рСО2 и рН крови поддерживаются нормальными, человек, как правило, не ощущает недостатка кислорода. Следствием этого могут быть опасные ситуации, возникающих в быту или при подключении человека к дыха! тельным аппаратам (замкнутым системам с газовыми смеся! ми). Наиболее распространенными являются отравления угарным газом (смерть в гараже, другие бытовые отравления), когда человек из!за отсутствия явных ощущений удушья не
предпринимает защитных действий.
Рецепторы дыхательных путей и легких. Особенно обильно снабжены рецепторами верхние дыхательные пути. В слизистой оболочке верхних носовых ходов между эпителиальными и опор! ными клетками расположены обонятельные рецепторы. Они представляют собой чувствительные нервные клетки, имеющие подвижные реснички, которые обеспечивают рецепцию пахучих веществ. Благодаря этим рецепторам и обонятельному анализа! тору организм получает возможность восприятия запахов, сигна!
369
лизирующих об окружающей обстановке, наличии пищевых ве! ществ, вредных агентов. Воздействие некоторых пахучих веществ вызывает рефлекторное изменение проходимости дыхательных путей (в частности, у людей с обструктивным бронхитом может вызвать астматический приступ).
Остальные рецепторы дыхательных путей и легких подраз! деляют на три группы: 1) рецепторы растяжения, 2) ирритант! ные, 3) юкстаальвеолярные.
Рецепторы растяжения располагаются в мышечном слое дыхательных путей. Адекватным раздражителем для них является растяжение мышечных волокон, обусловленное из! менением внутриплеврального давления и давления в просве! те дыхательных путей при осуществлении вдоха. Важнейшая функция этих рецепторов – контроль за степенью растяжения легких. Благодаря этим рецепторам функциональная система регуляции дыхания контролирует интенсивность вентиляции легких.
Имеется также ряд экспериментальных данных о наличии в легких рецепторов спадения, активирующихся при сильном уменьшении объема легких.
Ирритантные рецепторы обладают свойствами механо! и хеморецепторов. Они расположены в слизистой оболочке дыхательных путей и активируются при действии интенсивной струи воздуха во время вдоха или выдоха, действии крупных пылевых частиц, скоплении гноя, слизи, попадании в дыха! тельные пути частиц пищи. Эти рецепторы чувствительны так! же к действию раздражающих газов (аммиак, пары серы) и различных химических факторов.
Юкстаальвеолярные рецепторы расположены в интер! стициальном пространстве легочных альвеол у стенок крове! носных капилляров. Адекватным раздражителем для них явля! ется увеличение кровенаполнения легких и возрастание объ! ема межклеточной жидкости (они активируются, в частности, при отеке легких). Раздражение этих рецепторов рефлекторно
вызывает возникновение частого поверхностного дыхания.
Рефлекторные реакции с рецепторов дыхательных путей.
С рецепторов растяжения и ирритантных рецепторов возника! ют многочисленные рефлекторные реакции, обеспечивающие саморегуляцию дыхания, защитные рефлексы и рефлексы, влияющие на функции внутренних органов. Такое подразделе! ние этих рефлексов весьма условно, так как один и тот же реф!
370