

Полезные материалы за все 6 курсов / Ответы к занятиям, экзаменам / Лекции по общей биохимии
.pdfСтроение и обмен углеводов |
210 |
|
|
Все аминокислоты, кроме кетогенных лейцина и лизина, способны участвовать в синтезе глюкозы. Углеродные атомы глюкогенных аминокислот полностью включаются в молекулу глюкозы, других аминокислот (смешанных) – частично.
Кроме получения глюкозы, глюконеогенез обеспечивает и уборку "шлаков" – лактата, постоянно образуемого в эритроцитах или при мышечной работе и глицерола, являющегося продуктом липолиза в жировой ткани.
ОБХОД ДЕСЯТОЙ РЕАКЦИИ ГЛИКОЛИЗА
На этом этапе глюконеогенеза работают два основных фермента – в митохондриях пи-
руваткарбоксилаза и в цитозоле фосфоенолпируват-карбоксикиназа.
Вмитохондриях пируваткарбоксилаза превращает пировиноградную кислоту в оксалоацетат. Необходимо отметить, что эта реакция идет в клетке постоянно, являясь анаплеротической (пополняющей) реакцией ЦТК.
Далее оксалоацетат должен попасть в цитозоль и превратиться в фосфоенолпируват. Однако дело осложняется непроницаемостью мембраны для оксалоацетата. Зато через мембрану может пройти малат, предшественник оксалоацетата по ЦТК. Повернуть малатдегидрогеназ-
ную реакцию ЦТК вспять позволяет избыток НАДН в митохондриях. НАДН поступает из-окисления жирных кислот, активируемого в условиях недостаточности глюкозы в гепатоците. В результате малат накапливается, выходит в цитозоль и здесь превращается обратно в оксалоацетат.
Вцитоплазме фосфоенолпируват-карбоксикиназа осуществляет превращение оксалоацетата в фосфоенолпируват, для реакции требуется энергия ГТФ. От молекулы отщепляется тот же углерод, что и присоединяется.
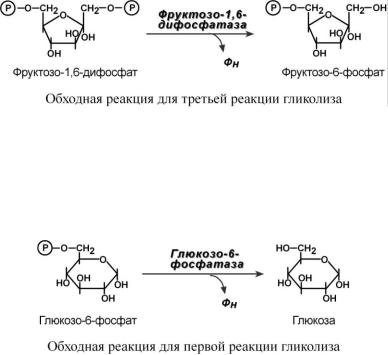
ОБХОД ТРЕТЬЕЙ РЕАКЦИИ ГЛИКОЛИЗА
Второе препятствие на пути синтеза глюкозы – фосфофруктокиназная реакция – преодолевается с помощью фермента фруктозо-1,6-дифосфатазы. Этот фермент есть в почках, печени, поперечно-полосатых мышцах. Таким образом, эти ткани способны синтезировать и фруктозо-6-фосфат и глюкозо-6-фосфат.
biokhimija.ru |
Тимин О.А. Лекции по общей биохимии (2020г) |
211 |
|
|
|
|
|
ОБХОД ПЕРВОЙ РЕАКЦИИ ГЛИКОЛИЗА
Последняя реакция катализируется глюкозо-6-фосфатазой. Она имеется только в печени и почках, следовательно, только эти ткани могут продуцировать свободную глюкозу. После завершения глюконеогенеза глюкоза выходит в кровь и разносится по организму.
В З А И М О О Т Н О Ш Е Н И Я Г Л И К О Л И З А И Г Л Ю К О Н Е О Г Е Н Е З А
Так как синтез и окисление глюкозы являются крайне важными для существования клетки (гликолиз) и всего организма (глюконеогенез), то регуляция этих процессов отвечает запросам органов и тканей при различных условиях.
Поскольку гликолитическое окисление глюкозы:
oявляется способом получения энергии и в аэробных и в анаэробных условиях, то оно непрерывно происходит во всех клетках и, конечно, должно и будет активировано при усилении работы клетки, например, сокращение миоцита, движение нейтрофилов;
oявляется источником глицерола и ацетил-SКоА, используемых для синтеза жиров в гепатоцитах и адипоцитах, то такое окисление активируется при избытке глюкозы в клетке. Глюконеогенез, как образование глюкозы в печени из неуглеводных компонентов, не-
обходим:
oпри гипогликемии во время мышечной нагрузки – синтез из молочной кислоты, поступающей из мышц, из глицерола, образующегося при мобилизации жиров;
oпри гипогликемии при голодании – синтез из аминокислот, образующихся при катаболизме белков, и из молочной кислоты.
Таким образом, при голодании или физической нагрузке глюконеогенез, идущий в пе-
чени, обеспечивает глюкозой все остальные клетки и органы (эритроциты, нервная ткань, мышцы и др.), в которых активны процессы, требующие глюкозу. Поступление глюкозы в эти клетки также необходимо, чтобы поддержать концентрацию оксалоацетата и обеспечить сгораниев ЦТК ацетил-SКоА, получаемого из жирных кислот или кетоновых тел.
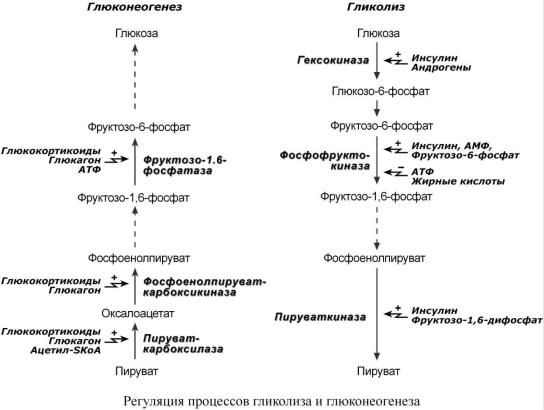
Р Е Г У Л Я Ц И Я Г Л И К О Л И З А И Г Л Ю К О Н Е О Г Е Н Е З А
В целом можно выделить два способа регуляции гликолиза и глюконеогенеза: гормональная и метаболическая, т.е. при помощи промежуточных или конечных продуктов обмена глюкозы.
Существуют три участка, на которых происходит регуляция: o первая реакция гликолиза,
o третья реакция гликолиза и обратимая ей,
Строение и обмен углеводов |
212 |
|
|
o десятая реакция гликолиза и обратимые ей.
Регуляция глюконеогенеза
Гормональная активация глюконеогенеза осуществляется глюкокортикоидами, кото-
рые увеличивают синтез пируват-карбоксилазы, фосфоенолпируваткарбоксикиназы,
фруктозо-1,6-дифосфатазы. Глюкагон стимулирует те же самые ферменты через аденилатциклазный механизм путем фосфорилирования.
Кроме гормонов, имеются метаболическая регуляция, при которой аллостерически ак-
тивируется пируваткарбоксилаза при помощи ацетил-SКоА, фруктозо-1,6-дифосфатаза
при участии АТФ.
Регуляция гликолиза
Гликолиз стимулируется инсулином, повышающим количество молекул гексокиназы, фосфофруктокиназы, пируваткиназы. В печени активность глюкокиназы регулируется гормонами: активацию вызывает инсулин и андрогены, подавляют ее активность глюкокортикоиды и эстрогены. Активность гексокиназ других клеток повышается инсулином, адреналином, тиреоидными гормонами, снижается – глюкокортикоидами и соматотропином.
К метаболической регуляции наиболее чувствительна фосфофруктокиназа. Она активируется АМФ и собственным субстратом, ингибируется – АТФ, лимонной кислотой, жирными кислотами. Пируваткиназа активируется фруктозо-1,6-дифосфатом. Гексокиназа клеток (кроме гепатоцитов) ингибируется продуктом собственной реакции – глюкозо-6-фосфа- том.
В К Л Ю Ч Е Н И Е Н Е У Г Л Е В О Д Н Ы Х В Е Щ Е С Т В В Г Л Ю К О Н Е О Г Е Н Е З
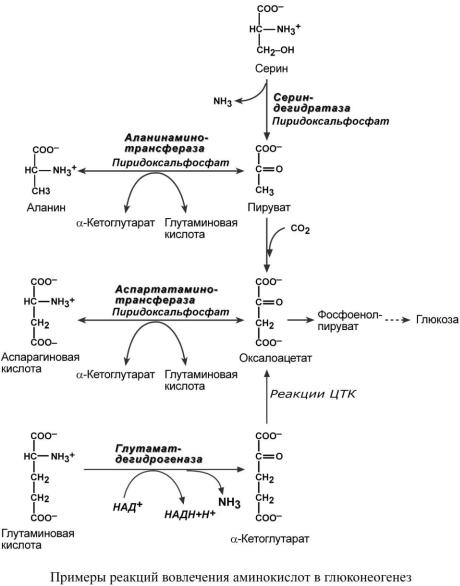
Синтез глюкозы из аминокислот
Ряд аминокислот являются глюкогенными, то есть их углеродные скелеты в той или иной степени способны включаться в состав глюкозы. Такими является большинство аминокислот, кроме лейцина и лизина, углероды которых никогда не участвуют в синтезе углеводов.
biokhimija.ru |
Тимин О.А. Лекции по общей биохимии (2020г) |
213 |
|
|
|
|
|
В качестве примера синтеза глюкозы из аминокислот рассмотрим участие в этом процессе глутамата, аспартата, серина и аланина.
Аспарагиновая кислота (после реакции трансаминирования) и глутаминовая кислота (после дезаминирования) превращаются в метаболиты ЦТК, соответственно, в оксалоацетат и-кетоглутарат.
Аланин, трансаминируясь, образует пировиноградную кислоту, которая способна карбоксилироваться до оксалоацетата. Оксалоацетат, являясь первым элементом в процессе глюконеогенеза, далее включается в состав глюкозы.
Серин в трехступенчатой реакции под воздействием сериндегидратазы теряет аминогруппу и превращается в пируват, который вступает в глюконеогенез.
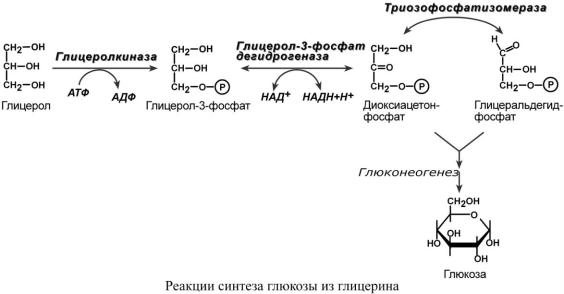
Синтез глюкозы из глицерина
При физической нагрузке под влиянием адреналина или при голодании, особенно длительном, под влиянием глюкагона и кортизола, в адипоцитах активно происходит распад триацилглицеролов. Одним из продуктов этого процесса является спирт глицерин, который поступает в печень. Здесь он фосфорилируется, окисляется до диоксиацетонфосфата и вовлекается в реакции глюконеогенеза.
Строение и обмен углеводов |
214 |
|
|
Синтез глюкозы из молочной кислоты
При физической нагрузке в мышцах продуцируется большое количество молочной кислоты. Также молочная кислота непрерывно образуется эритроцитами, независимо от состояния организма. С током крови она поступает в печень, где после проникновения в клетку превращается в пируват. Далее реакции идут по классической схеме.
Суммарная реакция синтеза глюкозы из молочной кислоты:
Лактат + 4АТФ + 2ГТФ + 2H2O Глюкоза + 4АДФ + 2ГДФ + 6Фн
Г Л Ю К О З О - Л А К Т А Т Н Ы Й И Г Л Ю К О З О - А Л А Н И Н О В Ы Й Ц И К Л Ы
Глюкозо-лактатный цикл (цикл Кори) – это связь глюконеогенеза в печени и образования лактата в эритроцитах или мышцах из глюкозы. В эритроцитах молочная кислота образуется непрерывно, так как для них анаэробный гликолиз является единственным способом образования энергии.
В скелетных мышцах высокое накопление молочной кислоты является следствием гликолиза при очень интенсивной, субмаксимальной мощности, работе. Но даже при работе низкой и средней интенсивности в скелетной мышце образуется некоторое количество лактата. Утилизоваться он может только одним способом – превратиться в пировиноградную кислоту. Однако, сама мышечная клетка ни при работе, ни во время отдыха не способна превратить лактат в пируват из-за особенностей изофермента ЛДГ-5.
Поэтому во время и после нагрузки (при восстановлении) лактат удаляется из мышцы. Это происходит довольно быстро, всего через 0,5-1,5 часа молочной кислоты в мышце уже нет. Малая часть молочной кислоты выводится с мочой. Большая часть лактата захватывается гепатоцитами, окисляется в пировиноградную кислоту и вступает на путь глюконеогенеза.
Полученная глюкоза используется самим гепатоцитом или выходит в кровь, пополняя запасы в мышце или других клетках.

biokhimija.ru |
Тимин О.А. Лекции по общей биохимии (2020г) |
215 |
|
|
|
|
|
Целью глюкозо-аланинового цикла также является уборка пирувата, но, кроме этого решается еще одна немаловажная задача – уборка аминного азота из мышцы. При мышечной работе и в покое в миоците распадаются белки и образуемые аминокислоты трансаминируются с -кетоглутаратом. Полученный глутамат взаимодействует с пируватом. Образующийся аланин является транспортной формой азота и пирувата из мышцы в печень. В гепатоците идет обратная реакция трансаминирования, аминогруппа передается на синтез мочевины, пируват используется для синтеза глюкозы.
Глюкоза, образованная в печени из лактата или аланина, возвращается обратно в мышцы, восстанавливая во время отдыха запасы гликогена. Также она может распределиться по другим органам.
Кроме мышечной работы, глюкозо-аланиновый цикл активируется во время голодания, когда мышечные белки распадаются и многие аминокислоты используются в качестве источника энергии, а их азот необходимо доставить в печень.
МЕ ТАБ ОЛИЗМ Э Т АН ОЛ А
Ворганизм поступают и в результате метаболизма образуются в клетках головного мозга
ипечени, в других тканях, при жизнедеятельности микрофлоры кишечника различные спирты (алифатической, ароматической, стероидной природы, ретиноиды, фарнезол и др.) и альдегиды (ароматические, алифатические, продукты пероксидации липидов и др.), которые могут являться промежуточными метаболитами или конечными продуктами. Взаимопревращение спиртов и альдегидов осуществляют алкогольдегидрогеназы. Их существует 6 классов, в каждом классе имеются многочисленные изоферменты, обнаруженные во многих тканях.
С медицинской и социальной точки зрения определенный интерес вызывает метаболизм этилового спирта в организме человека.
У Т И Л И З А Ц И Я Э Т А Н О Л А
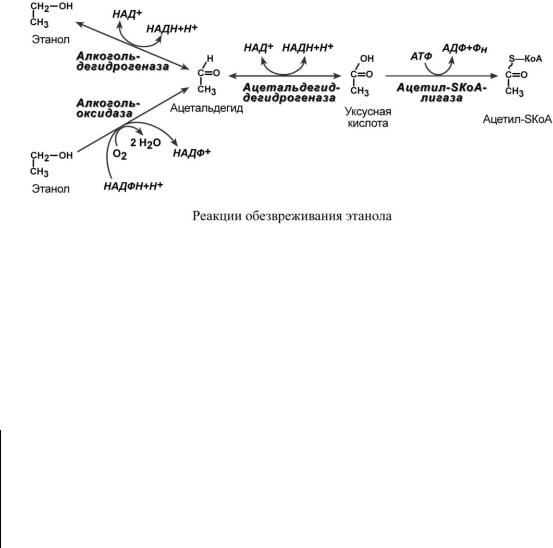
Метаболизм поступающего этанола в организме происходит в печени. Первый путь заключается в окислении спирта по алкогольдегидрогеназному пути до уксусной кислоты, которая в виде ацетил-SКоА поступает в ЦТК. Через этот путь проходит 80-90% всего этанола.
Строение и обмен углеводов |
216 |
|
|
За окисление оставшейся части этанола отвечает алкогольоксидаза (цитохром P450), также называемая микросомальная этанолокисляющая система (МЭОС) При регулярном поступлении этанола доля микросомального окисления возрастает, так как этанол является индуктором алкогольоксидазы и количество ее молекул увеличивается.
Поскольку при утилизации этанола образуется большое количество НАДН, то в цитозоле гепатоцитов активируется 11-я реакция гликолиза (превращение пирувата в лактат) и восстановление диоксиацетонфосфата в глицерол-3-фосфат. Все это приводит к гипогликемии, так как пировиноградная кислота и диоксиацетонфосфат являются субстратами глюконеогенеза.
Одновременно накопление "алкогольного" ацетил-SКоА ингибирует пируватдегидрогеназу, что еще больше усиливает накопление молочной кислоты и вызывает лактоацидоз.
Одновременно из-за относительной недостаточности оксалоацетата, использованного в глюконеогенезе, избыток "алкогольного" ацетил-SКоА не успевает окислиться в цикле трикарбоновых кислот и перенаправляется на синтез кетоновых тел, что обеспечивает возникно-
вение кетоацидоза.
Если запасы гликогена в печени изначально невелики (голодание, недоедание, астеническое телосложение) или израсходованы (после физической работы), то при приеме алкоголя натощак гипогликемия наступает быстрее и может быть причиной потери сознания. К этому стоит добавить сильный диуретический эффект этанола, ведущий к быстрому обезвоживанию организма и снижению кровоснабжения головного мозга со всеми вытекающими последствиями.
С П И Р Т О В О Е Б Р О Ж Е Н И Е
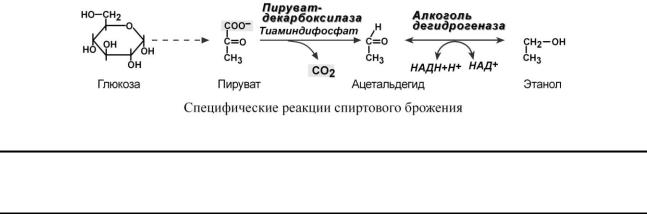
Образование этилового спирта из глюкозы происходит в анаэробных условиях в дрожжах и некоторых видах плесневых грибков. Суммарное уравнение реакции:
C6H12О6 2 CО2 + 2 С2Н5ОН + 2 АТФ
До стадии образования пирувата реакции спиртового брожения совпадают с реакциями гликолиза. Отличия заключаются только в дальнейшем превращении пировиноградной кислоты.
Цель этих превращений у указанных организмов – удалить пируват из клетки и окислить НАДН, который образовался в 6-й реакции.
biokhimija.ru |
Тимин О.А. Лекции по общей биохимии (2020г) |
217 |
|
|
|
|
|
ПЕ Н ТОЗОФО СФ АТНЫ Й П У ТЬ ОК ИСЛЕ Н ИЯ ГЛЮК ОЗЫ
Наиболее активно реакции пентозофосфатного пути (ПФП) идут в цитозоле клеток печени, жировой ткани, эритроцитах, коре надпочечников, молочной железе при лактации, в гораздо меньшей степени в скелетных мышцах. Этот путь окисления глюкозы не связан с образованием энергии, а обеспечивает анаболизм клеток. В связи с этим у новорожденных и детей первых лет жизни его активность довольно высока.
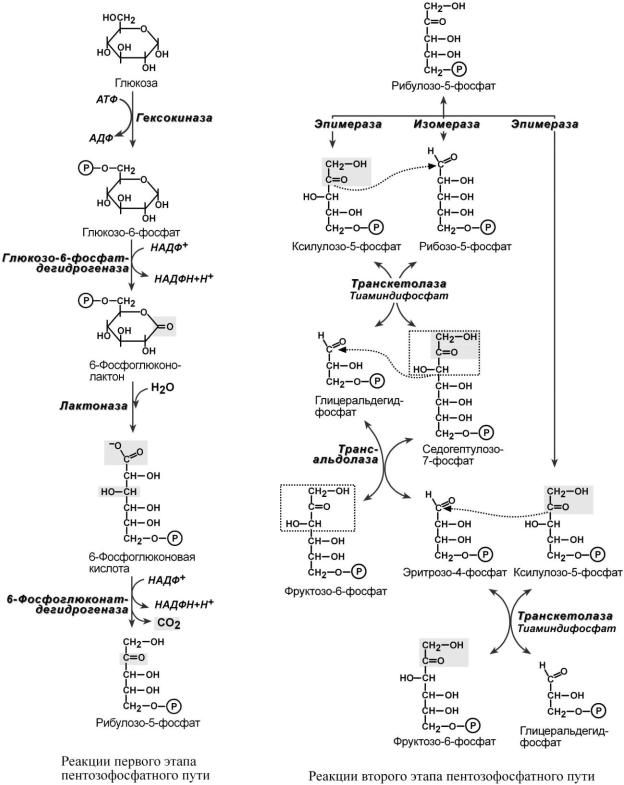
Пентозофосфатный путь (см стр.240) включает два этапа – окислительный и неокислительный.
На первом, окислительном, этапе глюкозо-6-фосфат в трех реакциях превращается в рибулозо-5-фосфат, реакции сопровождаются восстановлением двух молекул НАДФ до НАДФН.
На этом этапе происходит регуляция процесса: инсулин повышает активность глюкозо- 6-фосфат-дегидрогеназы и фосфоглюконат-дегидрогеназы.
Второй этап – этап структурных перестроек, благодаря которым пентозы способны возвращаться в фонд гексоз. В этих реакциях рибулозо-5-фосфат изомеризуется до рибозо-5- фосфата и ксилулозо-5-фосфата. Далее под влиянием ферментов транскетолазы и трансальдолазы происходят структурные перестройки с образованием других моносахаридов. При реализации всех реакций второго этапа пентозы превращаются во фруктозо-6-фосфат и глицеральдегидфосфат. Из глицеральдегид-3-фосфата при необходимости могут образоваться гексозы.
ЗН А Ч Е Н И Е П Е Н Т О З О Ф О С Ф А Т Н О Г О П У Т И
1.Значение первого этапа пентозофосфатного пути заключается в синтезе НАДФН. Образованный НАДФН используется:
o для синтеза жирных кислот,
o для синтеза холестерола и других стероидов (например, желчных кислот),
oдля синтеза глутаминовой кислоты из α-кетоглутаровой кислоты (реакция восстановительного аминирования),
o для синтеза дезоксирибонуклеотидов,
o для систем защиты клетки от свободно-радикального окисления (антиоксидантные системы),
o для деятельности цепи микросомального окисления (цитохром Р450),
oдля восстановления железа (III) метгемоглобина в железо (II) нормального гемоглобина,
oдля реактивации фолиевой кислоты и витамина К.
2.Значение второго этапа двояко:
oлибо в первой реакции образуется рибозо-5-фосфат, который используется при синтезе рибонуклеотидов и остальные пентозы стекаются в эту реакцию. Рибозо-5-фосфат необходим для синтеза пуриновых и пиримидиновых нуклеотидов и нуклеиновых кислот,
Строение и обмен углеводов |
218 |
|
|
o либо образуемые пентозо-5-фосфаты превращаются во фруктозо-6-фосфат и глицер–аль- дегид-3-фосфат и вливаются в путь гликолиза.
При определенных условиях разные клетки могут выключать или подключать второй неокислительный этап.
biokhimija.ru |
Тимин О.А. Лекции по общей биохимии (2020г) |
219 |
РОЛЬ ПЕНТОЗОФОСФАТНОГО ПУ ТИ В РАЗЛИЧНЫХ КЛЕТКАХ
Метаболизм глюкозо-6-фосфата в пентозофосфатном пути может идти по различным вариантам. Судьба метаболитов пентозофосфатного пути зависит от ситуации, потребностей клетки и ее типа.
Пример 1
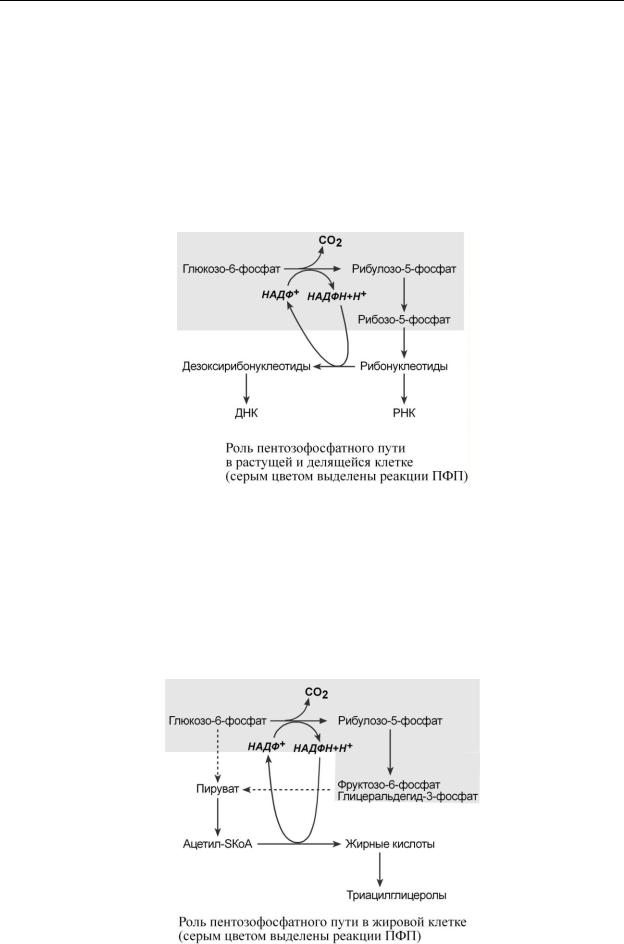
Рассмотрим ситуацию, когда потребность клетки в НАДФН и рибозо-5-фосфате сбалансирована. Например, при росте и делении клетки требуется рибоза для синтеза рибонуклеотидов и НАДФН для превращения рибонуклеотидов в дезоксирибонуклеотиды. При таких условиях реакции 1 этапа идут обычным порядком – образуется две молекулы НАДФН, но реакции 2 этапа обрываются на стадии синтеза рибозо-5-фосфата.
Пример 2
Если потребность в НАДФН значительно превышает потребность в рибозо-5-фосфате, как, например, в адипоцитах и печени при синтезе жирных кислот, то по окислительным реакциям пентозофосфатного пути образуются НАДФН и рибулозо-5-фосфат. Далее, под действием ферментов 2 этапа, рибулозо-5-фосфат превращается в пентозо-5-фосфаты и далее во фруктозо-6-фосфат и глицеральдегид-3-фосфат. Полученные продукты превращаются в пируват и ацетил-SKoA и, в основном, используются для синтеза жиров. Одновременно ГАФ может превращаться в глицерол-3-фосфат и уходить в биосинтез триацилглицеролов.