

Полезные материалы за все 6 курсов / Ответы к занятиям, экзаменам / Лекции по общей биохимии
.pdfСтроение и обмен нуклеотидов |
160 |
|
|

biokhimija.ru |
Тимин О.А. Лекции по общей биохимии (2020г) |
161 |
|
|
|
|
|
|
|
|
|
Н АР У ШЕ НИЕ ОБМ Е Н А ПИР ИМ ИД ИНОВЫ Х ОСНО В АН ИЙ
-АМИНОИЗОБУТИРАТАЦИДУРИЯ
Так как продукты катаболизма пиримидинов хорошо растворимы, то они хорошо выводятся с мочой и клинические симптомы патологий слабо выражены.
Примером подобного нарушения является -аминоизобутиратацидурия, при котором имеется дефект соответствующей трансаминазы. Заболевание протекает бессимптомно и встречается, в основном, у японцев и китайцев (около 25%).
ОР ОТАТАЦИДУРИЯ
Оротатацидурия (или оранжевая кристаллоурия) может быть наследственной и приобретенной.
Наследственная форма
Наследственная форма оротатацидурии весьма редка связана с одновременным дефек-
том оротатфосфорибозилтрансферазы и оротидилдекарбоксилазы, так как эти ферменты являются двумя активными центрами одного белка. Заболевание проявляется на втором полугодии жизни гипохромной мегалобластической анемией, лейкопенией, отставанием умственного и физического развития. Диагноз ставится при обнаружении в охлажденной моче оранжевых игольчатых кристаллов оротовой кислоты. Болезнь осложняется тем, что при недостатке УТФ и ЦТФ активируются первые реакции синтеза оротовой кислоты. При отсутствии лечения гибель наступает в первые годы жизни.
Однако, так как первопричиной заболевания является "пиримидиновый голод", его можно достаточно успешно корректировать с помощью приема уридина, при этом серьезно не опасаясь передозировки.
Приобретенная форма
Эта более частая форма оротатацидурии может наблюдаться:
oпри дефекте каких-либо ферментов синтеза мочевины, кроме карбамоилфосфат-синте- тазы. При этом карбамоилфосфат митохондрий (в норме используемый для образования мочевины) выходит из них и используется для избыточного синтеза оротовой кислоты.
Заболевание обычно сопровождается гипераммониемией.
oпри лечении подагры аллопуринолом, который превращается в оксипуринолмононуклеотид, являющийся ингибитором оротатдекарбоксилазы, что опять же ведет к накоплению оротата.
НУ К ЛЕ ОЗИД Ы И НУ К ЛЕ ОТ ИД Ы К АК ЛЕ К АР С ТВЕ Н НЫ Е ПР Е П АР АТЫ
Азотистое основание аллопуринол используется при лечении подагры, при мочекаменной болезни и синдроме Леша-Нихана.
Лечебный эффект ряда антивирусных препаратов основан на их строении – они являются производными азотистых оснований и нуклеозидов. Механизм действия различных нуклеозидов основан на их фосфорилировании в зараженных вирусом клетках с превращением в нуклеотиды, в конкуренции с обычными нуклеотидами за встраивание в вирусные нуклеиновые кислоты, что останавливает размножение вируса.

М АТРИЧНЫЕ БИОСИНТЕЗЫ
Основной фигурой матричных биосинтезов являются нуклеиновые кислоты. Они представляют собой полимерные молекулы, в состав которых входят азотистые основания пяти типов, пентозы двух типов и остатки фосфорной кислоты.
Азотистые основания в нуклеиновых кислотах могут быть пуриновыми (аденин, гуанин)
ипиримидиновыми (цитозин, урацил, тимин).
Взависимости от строения углевода выделяют рибонуклеиновые кислоты – содержат рибозу (РНК), и дезоксирибонуклеиновые кислоты – содержат дезоксирибозу (ДНК).
О С Н О В Н О Й П О С Т У Л А Т М О Л Е К У Л Я Р Н О Й Б И О Л О Г И И
В подавляющем большинстве случаев передача наследственной информации от материнской клетки к дочерней осуществляется при помощи ДНК (репликация). Для использования генетической информации самой клеткой необходимы РНК, образуемые на матрице ДНК (транскрипция). Далее РНК непосредственно участвуют на всех этапах синтеза белковых молекул (трансляция), обеспечивающих структуру и деятельность клетки.
На вышесказанном основана центральная догма молекулярной биологии, согласно которой перенос генетической информации осуществляется только от нуклеиновой кислоты (ДНК и РНК). Получателем информации может быть другая нуклеиновая кислота (ДНК или РНК) и белок.
С ТР ОЕ НИЕ НУ К ЛЕ И НОВ Ы Х К ИСЛОТ
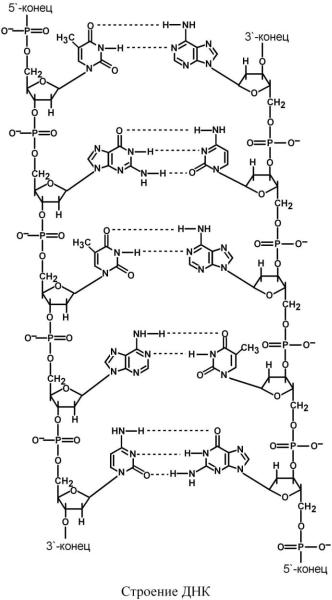
СТРОЕНИЕ ДЕЗОКСИРИБОН УКЛЕИНОВОЙ КИСЛОТЫ
ДНК (дезоксирибонуклеиновая кислота) представляет собой двухцепочечную молекулу, в каждой цепи мономерами выступают дезоксирибонуклеотиды – dАМФ, dГМФ, dЦМФ, dТМФ. Нуклеотиды связаны между собой через фосфатные мостики между 3' и 5' атомами углерода соседних дезоксирибоз. Сахарофосфатный остов в ДНК заряжен отрицательно благодаря заряду фосфатных групп. В то же время пуриновые и пиримидиновые основания гид-
рофобны.
Цепи ДНК обладают направленностью, т.е. имеют 3'-конец и 5'-конец, цепи антипараллельны, т.е. направлены в разные стороны. Имеется комплементарность азотистых оснований: аденин комплементарен тимину (А=Т), гуанин комплементарен цитозину (Г Ц).
biokhimija.ru |
Тимин О.А. Лекции по общей биохимии (2020г) |
163 |
|
|
|
|
|
СТРОЕНИЕ ХРОМАТИНА
ДНК – наиболее важная часть хромосом: две двухцепочечные молекулы ДНК образуют одну хромосому. Наиболее хорошо хромосомы видны перед митозом и во время него. В покоящихся клетках хромосомный материал выглядит нечетко и распределен по всему ядру. В таком состоянии он получил название "хроматин". В составе хроматина выделяют 60% белка (гистоны и кислые белки), 35% ДНК и около 5% РНК.
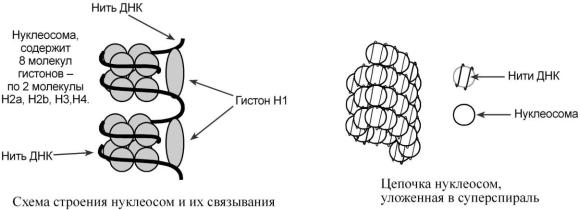
Хроматин уложен в виде сферических частиц – нуклеосом, соединенных друг с другом нитью ДНК. Нуклеосома представляет собой комплекс участка молекулы ДНК и восьми молекул гистонов. В составе нуклеосомы находятся по 2 молекулы гистонов Н2 , Н2 , Н3, Н4. Нить ДНК, последовательно контактируя с гистонами Н2 , Н2 , Н4, Н3, Н3, Н4, Н2, Н2, наматывается на гистоновое ядро, которое "маскирует" 146 пар оснований ДНК. Гистон Н1 связывается с нуклеосомой на участке входа и выхода ДНК, "склеивая" 2 оборота и "маскируя" еще 20 пар оснований. Таким образом, с помощью одной нуклеосомы замаскировано 166 пар оснований.
Кроме нуклеосом, в ядре присутствуют еще 2 уровня укладки ДНК: фибриллы диаметром 10 нм, состоящие из цепочки нуклеосом, и волокна, диаметром 30 нм, образующиеся при закручивании фибрилл в суперспираль. На виток такой спирали приходится 6-7 нуклеосом.
Матричные биосинтезы |
164 |
|
|
Участок ДНК между нуклеосомами называется спейсерным (англ: space – пространство), его длина варьирует в зависимости от вида организма и типа клеток. У человека она составляет около 50 пар нуклеотидов.
Благодаря наличию нуклеосом достигается уменьшение размеров хромосомы в 7 раз, далее происходит укладка в суперспираль фибриллы и „суперсуперспираль" волокна.
СТРОЕНИЕ РИБОНУКЛЕИНОВЫХ КИСЛОТ
Рибонуклеиновая кислота (РНК) представляет собой последовательность рибонуклеозидмонофосфатов – АМФ, ГМФ, ЦМФ, УМФ, связанных друг с другом 5'-3'-фосфодиэфир- ными связями. По строению РНК отличается от ДНК однонитевой структурой, значительно меньшей молекулярной массой, наличием урацила вместо тимина и рибозы вместо дезоксирибозы.
В клетке присутствует четыре типа РНК:
Матричные РНК (мРНК) представляют собой линейную последовательность нуклеотидов. К 5'-концу молекулы присоединен метилгуанозиндифосфат, на 3'-конце имеется полиадениловая последовательность (см ниже). Их функция – информационная, т.е. перенос информации о структуре белков от ДНК к месту их синтеза.
Малые РНК используются для созревания мРНК и некоторых других клеточных процессов.
Рибосомальные РНК (рРНК) прокариот и эукариот различны и отличаются величиной седиментации (скорости оседания молекулы при центрифугировании). Они участвуют в построении рибосом.
У прокариот есть три разновидности рРНК: 5S, 16S и 23S. Малую (30S) субчастицу рибосом образуют белки и 16S-рРНК; большую (50S) субъединицу – белки, 23S-рРНК и 5S-рРНК. У эукариот – в составе рибосом четыре разновидности рРНК: 5S, 5,8S, 18S и 28S. Малую (40S) субчастицу образуют белки и 18S-рРНК, большую (60S) – белки и 5S-, 5,8S-, 28S-рРНК.
biokhimija.ru |
Тимин О.А. Лекции по общей биохимии (2020г) |
165 |
|
|
|
|
|
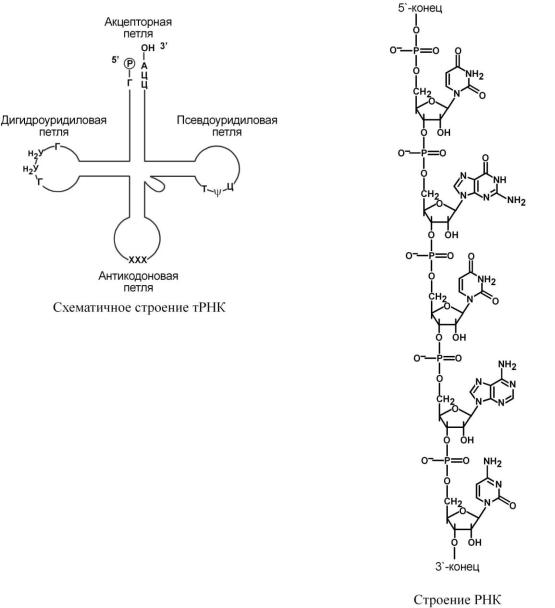
Транспортные РНК (тРНК) бактерий и эукариот включают 73-93 нуклеотида. Они переносят аминокислоты из цитозоля к рибосомам. На 5'-конце тРНК находится гуаниловый нуклеотид, на 3'-конце – триплет Ц-Ц-А. Вторичная структура тРНК напоминает клеверный лист, а третичная – латинскую букву L. В "клеверном листе" выделяют четыре участка (или ветви, петли), каждый из которых имеет собственную функцию:
o антикодоновый – соединяется с кодоном матричной РНК в рибосоме, o псевдоуридиловый – отвечает за связывание с рибосомой,
o дигидроуридиловый – отвечает за связывание с аминоацил-тРНК-синтазой, o акцепторный – связывает переносимую аминокислоту.
Матричные биосинтезы |
166 |
|
|
|
|
СИН ТЕ З НУ К ЛЕ И НОВЫ Х К И СЛО Т
Р Е П Л И К А Ц И Я Д Н К
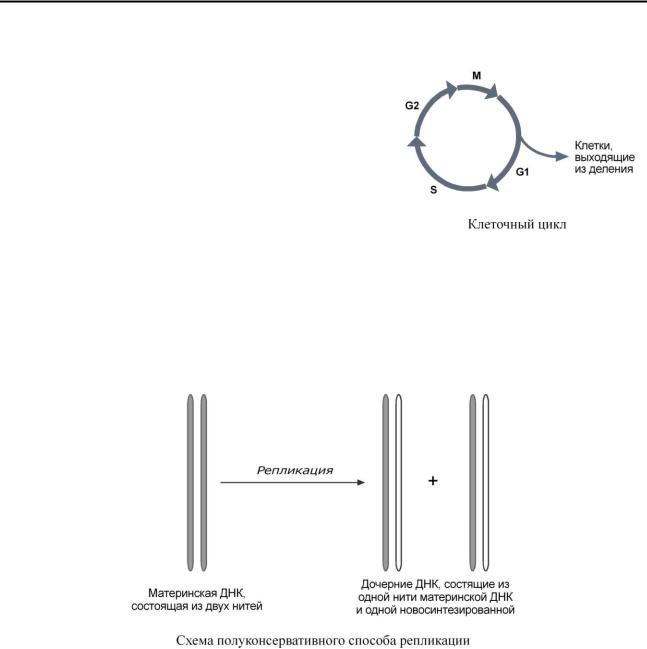
Синтез ДНК происходит не беспорядочно, а в строго определенный период жизни клетки. Всего выделяют 4 фазы клеточного цикла: митоз (М), синтетическую (S), пресинтетическую (G1, от англ. gap – интервал), постсинтетическую (G2).
Важное участие в регуляции смены фаз клеточного цикла занимают циклины – белки массой 35-90 кДа, уровень которых меняется в ходе клеточного цикла. По функции циклины – это активаторные субъединицы ферментов циклин-зависимых киназ (ЦЗК). Активные комплексы циклин-ЦЗК фосфорили-
руют внутриклеточные белки, изменяя их активность. Этим обеспечивается продвижение по клеточному циклу.
Синтез (репликация, удвоение) ДНК происходит в S-фазу клеточного цикла, когда клетка готовится к делению. Механизм репликации, как установили Мэтью Мезельсон и Франклин Сталь в 1957 г, полуконсервативный, т.е. на каждой нити материнской ДНК синтезируется дочерняя копия.
Как любой матричный биосинтез, репликация требует наличия нескольких компонентов: o матрица – в ее роли выступает материнская нить ДНК,
o растущая цепь – дочерняя нить ДНК,
o субстраты для синтеза – dАТФ, dГТФ, dЦТФ, ТТФ, o источник энергии – dАТФ, dГТФ, dЦТФ, ТТФ,
o ферменты.
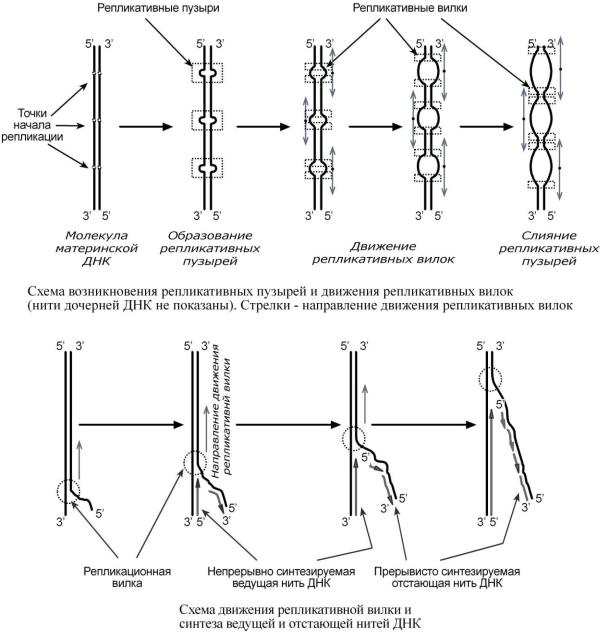
Синтез ДНК начинается в определенных участках, получивших название точка ori (англ. origin – начало). На каждой ДНК млекопитающих точек ori насчитывается около 100. Репликация распространяется от этих участков в обе стороны по нитям ДНК с образованием репли-
кативных "пузырей". В каждом таком "пузыре" имеются две репликативные "вилки", в
которых происходит расплетание, раскручивание и непосредственный синтез ДНК. При этом репликативные вилки удаляются друг от друга. В целом вся репликация ДНК у эукариот заканчивается за 9 часов.
biokhimija.ru |
Тимин О.А. Лекции по общей биохимии (2020г) |
167 |
|
|
|
|
|
Синтез новой цепи ДНК идет в направлении от 5'-конца к 3'-концу, т.е. 5'-конец новой ДНК остается свободным, следующие нуклеотиды присоединяются к 3'-гидроксильной группе предыдущего нуклеотида.
В репликативной вилке в направлении 5' 3' непрерывно (т.е. обычным присоединением последующих нуклеотидов к предыдущим через С3 и С5) синтезируется только одна нить, а именно та, для которой направление синтеза совпадает с направлением движения репликативной вилки и соответствует направлению материнской нити 3' 5'. По мере расплетания ДНК и движения репликативной вилки на этой материнской нити открываются участки,
где возможно безостановочное удлинение ведущей дочерней нити.
Направление 5' 3' для другой дочерней нити ДНК противоположно движению репликативной вилки. Поэтому синтез этой отстающей нити (в направлении 5' 3') возможен только после расплетания части ДНК и освобождения участка для синтеза.
Таким образом, синтез дочерней ДНК идет фрагментарно. По имени японского исследователя синтезируемые на отстающей цепи отрезки ДНК назвали фрагменты Оказаки.
Матричные биосинтезы |
168 |
|
|
В хронологическом порядке события репликации развертываются примерно следующим образом:
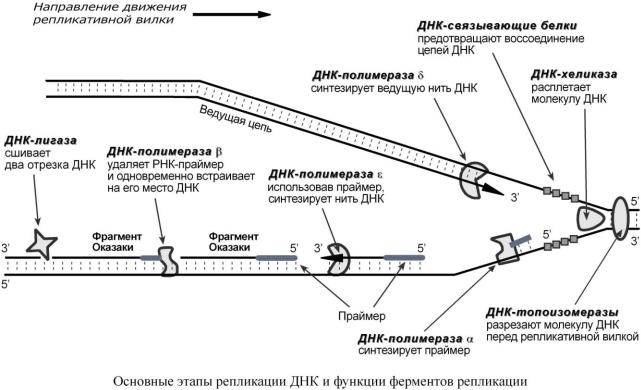
1.ДНК-топоизомеразы, находясь перед репликативной вилкой, разрезают молекулу ДНК для облегчения ее расплетания и раскручивания.
2.ДНК-хеликазы, следуя за топоизомеразами, раскручивают и расплетают молекулу ДНК.
3.ДНК-связывающие белки (ДСБ) связывают расплетенные нити ДНК и стабилизируют их, не допуская обратного "слипания" друг с другом.
4.ДНК-полимераза (греч.: – дельта), согласовано со скоростью движения репликативной вилки, осуществляет элонгацию ведущей цепи дочерней ДНК в направлении 5' 3' на матрице одной из нитей материнской ДНК (скорость 100 пар нуклеотидов в секунду).
5.ДНК-полимераза α присоединяется к другой нити ДНК сразу после расплетания и в направлении 5' 3' синтезирует праймер (РНК-затравку) – последовательность РНК на матрице ДНК длиной от 10 до 200 нуклеотидов. После этого фермент удаляется с нити ДНК.
6.ДНК-полимераза проводит синтез фрагмента (длина 150-200 нуклеотидов) отстающей цепи дочерней ДНК в продолжение праймера. Она работает до тех пор, пока не встретит праймер предыдущего фрагмента Оказаки (синтезированного ранее). После этого данный фермент удаляется с цепи.
7.ДНК-полимераза встает вместо ДНК-полимеразы , движется в том же направлении (5' 3') и удаляет рибонуклеотиды праймера, одновременно встраивая дезоксирибонуклеотиды на их место. Фермент работает до полного удаления праймера, после чего сходит с цепи.
8.ДНК-лигаза производит сшивку отрезка, синтезированного ДНК-полимеразой , и участка отстающей цепи дочерней ДНК, встроенного ДНК-полимеразой , (фрагментов Ока-
заки).

biokhimija.ru |
Тимин О.А. Лекции по общей биохимии (2020г) |
169 |
РЕГУЛЯЦИЯ РЕПЛИКАЦИИ
Лекарственная регуляция
1.Дауномицин и актиномицин D интеркалируют, т.е. встраиваются между нитей ДНК
ипрепятствуют продвижению ферментов репликации.
2.Новобиоцин снижает активность ДНК-гиразы у бактерий, подавляя их размножение.
3.Мелфалан и циклофосфамид превращаются в организме в соединения, которые вызывают присоединение группы CH3 или C2H5 к атому N7 гуанозина обеих цепочек молекулы ДНК, что приводит к перекрестной сшивке двух спиралей ДНК и остановке репликации.
П О В Р Е Ж Д Е Н И Я И Р Е П А Р А Ц И Я Д Н К
Так как на геном любой неделящейся клетки постоянно оказывает влияние окружающая среда, то вполне вероятны повреждения в составе нуклеотида, также возможно встраивание неправильного нуклеотида при репликации. Такие нарушения быстро определяются специальными ферментами, пораженный участок удаляется экзонуклеазами, заполняется
ДНК-полимеразой и сшивается ДНК-лигазой.
В случае изменения структуры основания (например, его дезаминирование) это основание удаляется ДНК-N-гликозидазой, затем другими ферментами удаляется дезоксирибоза и на ее место ДНК-полимеразой и ДНК-лигазой встраивается нужный нуклеотид.
Г И Б Р И Д И З А Ц И Я Д Н К - Д Н К И Д Н К - Р Н К
Если нагреть раствор ДНК выше температуры 90 С или сдвинуть рН в резко щелочную или резко кислую стороны, то водородные связи между нитями ДНК разрушаются и двойная спираль расплетается. Происходит денатурация ДНК или, по-другому, плавление. Если удалить агрессивный фактор, то происходит ренатурация или отжиг. При отжиге нити ДНК "отыскивают" комплементарные участки друг у друга и, в конце концов, вновь сворачиваются в двойную спираль.
Если в одной "пробирке" провести плавление и отжиг смеси ДНК человека и мыши, то некоторые участки цепей ДНК мыши будут воссоединяться с комплементарными участками цепей ДНК человека с образованием гибридов. Число таких участков зависит от степени родства видов. Чем ближе виды между собой, тем больше участков комплементарности нитей ДНК. Это явление называется гибридизация ДНК-ДНК.
Если в растворе присутствует РНК, то можно осуществить гибридизацию ДНК-РНК. Такая гибридизация помогает установить близость определенных последовательностей ДНК с какой-либо РНК.
Гибридизация ДНК-ДНК и ДНК-РНК используется как эффективное средство в молекулярной генетике.
С И Н Т Е З Р Н К – Т Р А Н С К Р И П Ц И Я
Прежде чем начнут синтезироваться белки, информацию об их строении необходимо "достать" из ДНК и доставить ее к месту синтеза белков. Этим занимаются информационные или матричные РНК. Одновременно клетке нужны транспортеры аминокислот – транспортные РНК и структурные компоненты органелл, синтезирующих белок, – рибосомальные РНК. Вся информация о строении транспортных и рибосомальных РНК также находится в ДНК.
Поэтому существует процесс переписывания или транскрипции данных с ДНК на РНК (англ. transcription – переписывание) – биосинтез РНК на матрице ДНК.
Как в любом матричном биосинтезе в транскрипции выделяют 5 необходимых элемен-
тов:
o матрица – одна из цепей ДНК, o растущая цепь – РНК,