

1.2.2. В'язкість крові
Кров являє собою приклад складної за своїм вмістом рідини. Вона є суспензією форменних елементів (еритроцитів, лейкоцитів, тромбоцитів) у водному колоїдному розчині - плазмі, сумарна концентрація білків у якій становить 6-9%. Експеримент виявив суттєву залежність в'язкості крові від її складу, що визначається показником гематокриту Не (мал. 1.9а), який дорівнює відношенню об'єму форменних елементів Vф до об'єму плазми крові Vпл:
 (1.11)
(1.11)
Оскільки об'єм форменних елементів в основному припадає на еритроцити, показник гематокриту характеризує вміст еритроцитів у крові.
Як свідчить наведена на малюнку залежність ηвідн = = f(Не), в'язкість крові змінюється у досить широкому діапазоні по відношенню до норми (N). Вона зростає при поліцитемії і зменшується при анемії.
Відомо декілька емпіричних формул, що зв'язують коефіцієнт в'язкості крові з показником гематокриту:
1
![]()
![]() (1.12)
(1.12)
де ηо - в'язкість плазми, α, β, γ- емпіричні константи, значення яких залежить від концентрації та форми суспен-зованих елементів.
Дослідження залежності в'язкості крові від швидкості деформації зсуву (градієнта швидкості) свідчать про те, що кров не є ньютонівською рідиною. При великих градієнтах швидкості (наприклад, в артеріальних судинах) в'язкість крові наближається до в'язкості води, у той час як при малих значеннях швидкості деформації зсуву в'язкість у п'ять і більше разів перевищує в'язкість води (мал. 1.9б).
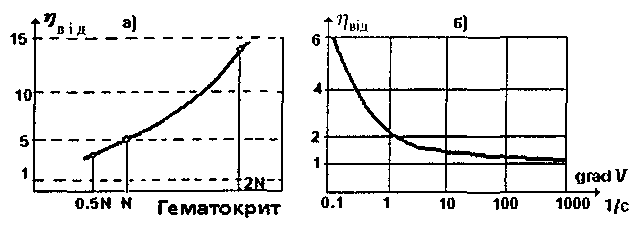
Мал. 1.9. Зміна в'язкості крові при зміні: а) форменного складу крові, б) швидкості деформації зсуву.
Величина відносної в'язкості крові може бути використана у діагностиці захворювань (див. табл. 1.1). Залежність коефіцієнта в'язкості від градієнта швидкості dυ/dy обумовлена здатністю еритроцитів до агрегації - утворенню "монетних стовпчиків" та їх конгломератів. Із збільшенням градієнта швидкості стовпчики руйнуються, і коефіцієнт в'язкості зменшується внаслідок дезагрегації та деформації еритроцитів.
Таблиця 1.1.
|
Відносна в'язкість крові |
Результат |
|
4.2-6.0 |
Норма |
|
<2.0 |
Анемія |
|
>10.0 |
Поліцитимія |
Зменшення в'язкості крові при її переході з венозного русла в артеріальне фізіологічне виправдане. У цьому випадку значно зменшуються витрати м'язової енергії міокарду на просування крові вздовж артеріального русла, в якому величини швидкостей деформації зсуву (а отже і сили внутрішнього тертя) досить значні (вони у сотні разів перебільшують значення останніх у венозній ділянці судинної системи).
1.2.3. В'язко-пружні властивості біологічних тканин
Біологічні структури (м'язи, судини, сухожилля, тканини легенів, шкіра тощо) являють собою в'язко-пружні системи, їх поведінка вивчається на моделях, що вміщують пружні (Е) та в'язкі (η) елементи, у деяких випадках до них додають і елементи зовнішнього тертя (К).

Мал. 1.10. Механічні моделі тканин: 1) пружний елемент; 2) в'язкий елемент; 3) елемент внутрішнього тертя; 4) послідовне з'єднання в'язкого та пружного елементів; 5) паралельне з'єднання в'язкого та пружного елементів.
Пружний елемент являє собою ідеальну пружину, для якої виконується закон Гука. В'язкий елемент може бути поданий у вигляді циліндра, який заповнений в'язкою рідиною з нещільним поршнем. Для витягування поршня необхідно прикласти деяку зовнішню силу, яка компенсує сили в'язкого тертя, що виникають при плині рідини крізь зазор.
Напруження, що створюються цими елементами під дією зовнішніх сил, дорівнюють:
 для
пружного
елементу;
для
пружного
елементу; для
в’язкого елементу;
для
в’язкого елементу;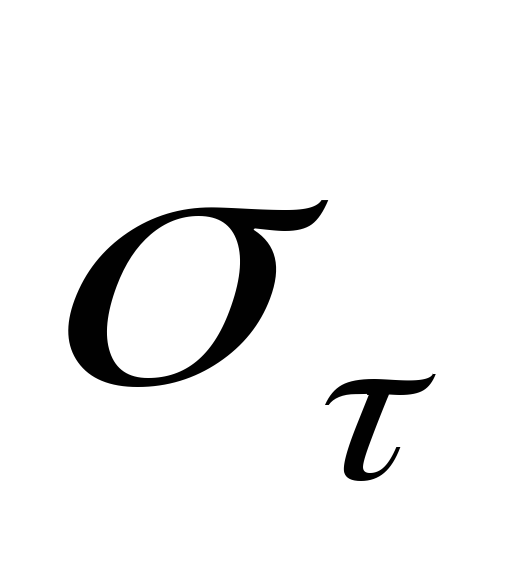 ~
KFn
для
елементу зовнішнього тертя при силі
нормального тиску Fn
і
коефіцієнті тертя К.
~
KFn
для
елементу зовнішнього тертя при силі
нормального тиску Fn
і
коефіцієнті тертя К.
Для відтворення механічних властивостей біологічних тканин використовують моделі, що складаються з цих елементів. Найпростішими моделями є тіло Максвелла і тіло Фойгта, що являють собою послідовне і паралельне з'єднання пружного та в'язкого елементів (див. мал. 1.10). Ці моделі дозволяють відтворити такі динамічні властивості тканин, як повзучість та релаксація напруження.
Повзучість - це явище зміни з часом розмірів зразка в умовах дії постійного напруження. Якщо у біологічних тканинах швидко створити, а потім підтримувати постійним деяке напруження, то з часом відбувається поступове подовження зразка аж до розриву тканин, навіть при умові, що постійне напруження має менше значення, ніж межа міцності матеріалу. Динаміку повзучості подано на мал. 1.1 la. Зміна розмірів відбувається тим швидше, чим більше напруження, що підтримується у зразку (порівняйте криві 1, 2 та 3, для яких (σ1 > σ2 > σ3).

Мал. 1.11. Динамічні властивості біологічних тканин: а) повзучість - зміна деформації тіла за умови постійного напруження σ (σ1 > σ2 > σ3); б) релаксація напруження - зменшення σ в умовах постійної деформації.
Релаксація напруження - явище зменшення з часом величини напруження у зразку при підтримці постійної величини деформації. Якщо швидко розтягнути зразок і, підтримуючи постійною отриману деформацію, вимірювати напруження в ньому протягом деякого часу, помітним стане його зменшення з часом (мал. 1.116). Пунктирними лініями на обох мал. 1.11 відтворено поведінку чисто пружних тіл. Релаксація напруження і повзучість суто динамічні процеси - час їх існування вимірюється секундами або хвилинами. Наприклад, для м'язів час зменшення напруження на 40% становить близько 10 секунд.
Ці процеси легко пояснюються механічними моделями, наведеними на мал. 1.10 (позиція 4). Спочатку під дією зовнішніх сил деформується пружний елемент, а потім починається "плин" в'язкого елементу, змінюється його розмір, що викликає зміну як довжини, так і напруження. За допомогою моделі Максвелла легко отримати експоненціальний закон релаксації напруження
![]() ,
(1.13)
,
(1.13)
де α- постійна часу релаксації, σ0 = Еε- початкове напруження.
Явище повзучості також можна описати експоненціальним законом
![]() (1.14)
(1.14)
де ε0= σ0/Е - початкова деформація, τ- характерний час процесу повзучості, що дорівнює відношенню коефіцієнта в'язкості і модуля Юнга. Формулу τ= η/Е легко отримати з міркувань розмірності. Дійсно, розмірність η є Па*с, a розмірність Е дорівнює Н/м2 = Па. Тому єдина комбінація величин ηі Е, що має розмірність часу [τ] = с, є їх відношення η/е.
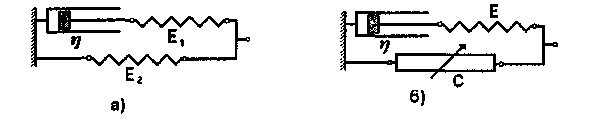
Мал. 1.12. Приклади механічних моделей біологічних тканин: а) трьохелементна модель для дослідження механічних властивостей 1в'язко-пружних тканин; б) трьохелементна модель м'язів, що включає скорочувальний елемент С.
Модель Фойгта дозволяє пояснити поступове зростання з часом напруження у зразку, якщо до тіла прикласти зусилля, що змінюється стрибкоподібне. Динаміка експоненціального зменшення напруження чи деформації відрізняється від експерименту. Кращих результатів можна досягти, якщо розглядати моделі, що включають до себе кілька пружних та в'язких елементів. Приклад однієї з таких моделей наведено на мал. 1.12. На цьому ж малюнку наведено одну з найпростіших моделей м'язів, що включає скорочувальний елемент С, котрий являє собою механохімічний конвертор, який перетворює енергію хімічних реакцій на механічну енергію. Ця механічна енергія витрачається на створення напруження і здійснення роботи по скороченню м'язів.
Наявність в'язко-пружних елементів, з'єднаних зі скорочувальними елементами, забезпечують ті гнучкі функціональні властивості, які притаманні цілому ряду фізіологічних систем (наприклад, серцево-судинній, м'язовій і ряду інших) для виконання призначених їм функцій в умовах зміни як властивостей самої системи, так і зовнішних навантажень. Це явище притаманне системам, які адаптуються. Так, зміна тонусу судин еласто-м'язового типу дозволяє реалізувати таке явище, як депонування крові, при якому значне збільшення об'єму судини можливе лише при повній релаксації стінки судини і зменшенні її пружності. Навпаки, при необхідності вигнання крові з депо об'єм судин зменшується, релаксація напруження
відбувається при інших розмірах судини, у цьому випадку зростає і модуль об'ємної пружності судин (їх тонус).