

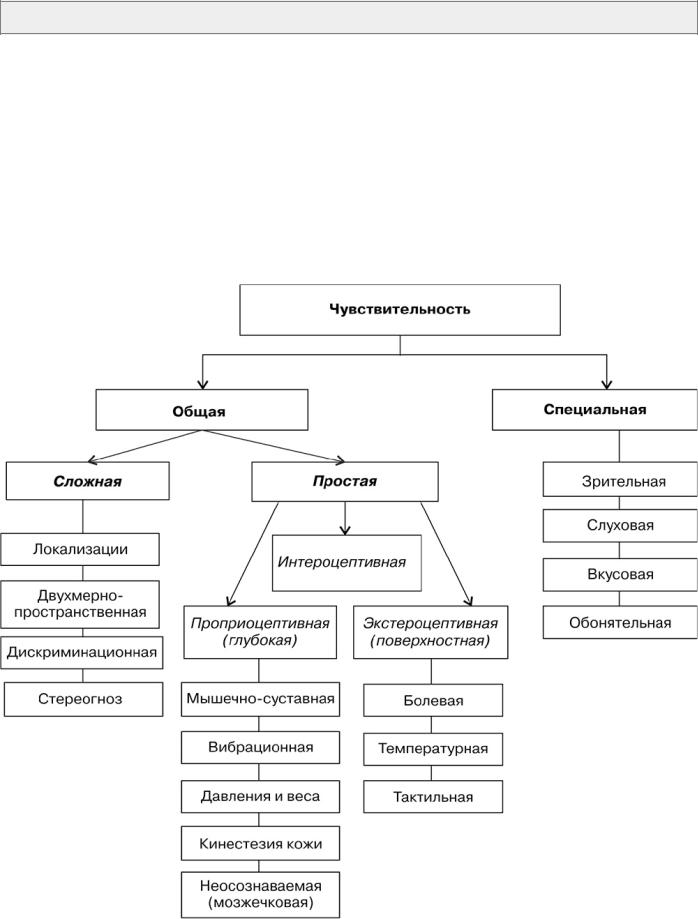
3.1. Виды чувствительности
Существуют различные варианты систематизации видов чувствительности. Достаточно распространена классификация по месту возникновения раздражения: экстероцептивная, проприоцептивная, интероцептивная.
В последние десятилетия вновь повысился интерес к систематизации, основанной на филогенетическом возрасте: протопатическая, ноцицептивная, таламическая. В повседневной клинической практике наиболее распространена описательная классификация. Один из возможных вариантов классификации видов чувствительности представлен на рис. 1.
Рис. 1. Клиническая классификация видов чувствительности
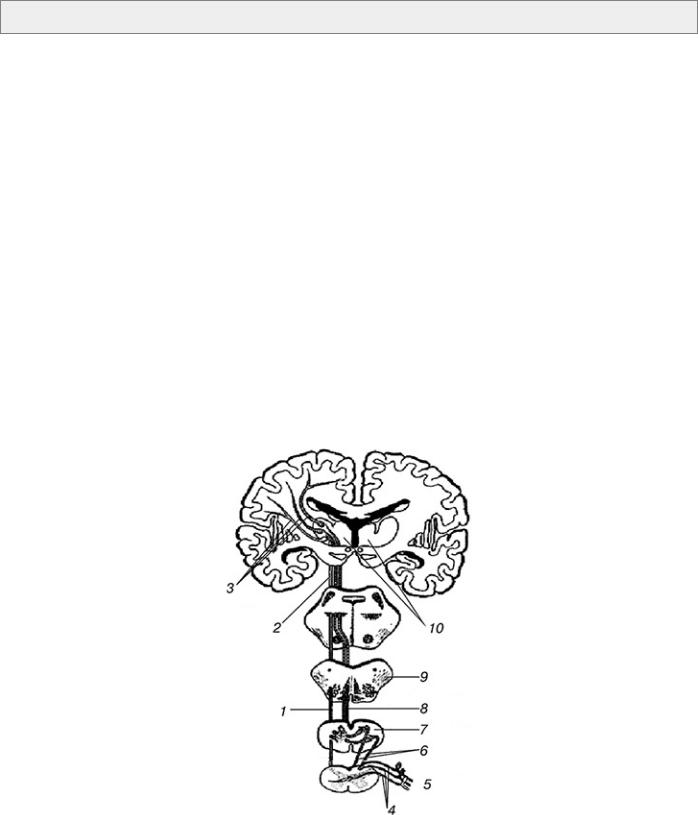
3.2. Проводники чувствительности
Проводники поверхностной чувствительности, как и проводники глубокой чувствительности, образованы тремя нейронами.
Тела первых нейронов пути поверхностной чувствительности (как и пути глубокой чувствительности) расположены в спинномозговых ганглиях (рис. 2). Периферические отростки (дендриты) первых нейронов пути поверхностной чувствительности в составе сплетений и периферических нервов достигают специфических рецепторов и соответствующих дерматомов.
Нервные волокна различны как по морфологии, так и по скорости проведения импульса. Толстые миелиновые волокна проводят импульс со скоростью 50—100 м/ с, тонкие миелиновые – 10–20 м/с, безмиелиновые – 0,5–2 м/с. Они различаются и по филогенетическому возрасту: наиболее древние – безмиелиновые, наиболее молодые – толстые миелиновые. Это важно для понимания характера возникающих нарушений, так как существует общее филогенетическое правило: молодые структуры (волокна) подвержены воздействию повреждающих факторов в значительно большей степени, чем филогенетически древние образования.
Рис. 2. Схема хода волокон путей глубокой и поверхностной чувствительности:
1 – спиноталамический путь; 2 – медиальная петля; 3 – таламо-корковый путь; 4 – задний корешок спинного мозга; 5 – спинномозговой ганглий (первый нейрон); 6
– пучки Голля и Бурдаха; 7 – ядра задних столбов (второй нейрон); 8 – бульботаламический путь; 9 – межоливный слой; 10 – таламус (третий нейрон)
Аксоны псевдоуниполярных нейронов спинномозговых ганглиев в составе
задних корешков, насчитывающих более 800 тыс. волокон, достигают спинного мозга. Далее волокна болевой, температурной и часть волокон тактильной чувствительности проникают в задние рога, проходя по заднелатеральному пути (краевая зона Лиссауэра) и через студенистое вещество, отдают коллатеральную ветвь к нейронам передних рогов (для соответствующего сегментарного рефлекса) и
вступают в контакт с отростками вторых нейронов пути поверхностной чувствительности, расположенных в собственных ядрах задних рогов спинного мозга.
Аксоны вторых нейронов пути поверхностной чувствительности через переднюю серую спайку (наискось и вверх на 2–3 сегмента) переходят на противоположную сторону, формируют латеральный спиноталамический путь (в боковом канатике) и передний спиноталамический путь (в переднем канатике). Латеральный спиноталамический путь проводит болевую и температурную чувствительность, передний спиноталамический путь – тактильную чувствительность. Тактильную чувствительность от кожи промежности передает центральный спиноталамический путь, располагающийся вокруг центрального канала. Важно отметить, что часть путей тактильной чувствительности проходит в задних канатиках совместно с проводниками глубокой чувствительности.
Следует иметь в виду особенность расположения волокон бокового и переднего спиноталамических путей: наиболее латерально располагаются волокна от дерматомов ноги, наиболее медиально – от дерматомов руки. Этот закон эксцентрического расположения волокон (закон Ауэрбаха – Флатау) важен для топической диагностики: при экстрамедуллярном процессе нарушения поверхностной чувствительности будут восходящего типа, при интрамедуллярном – нисходящего.
В составе бокового, переднего и центрального спиноталамических путей волокна поднимаются вверх и на уровне продолговатого мозга образуют единый спиноталамический путь, который в анатомии называют спинномозговой петлей (lemniscus spinalis). Этот путь проходит продолговатый мозг, мост, ножки мозга и заканчивается у вентролатеральных ядер таламуса.
Тела третьих нейронов пути поверхностной чувствительности (как и пути глубокой чувствительности) расположены в вентролатеральных ядрах таламуса. Их аксоны проходят через заднюю ножку внутренней капсулы, веерообразно расходятся в белом веществе (через лучистый венец) и достигают коры в постцентральной извилине и прилежащих участках теменной доли. В постцентральной извилине существует четкая соматотопическая проекция проводников по отношению к определенным частям тела (сенсорный гомункулус Пенфилда): в верхних отделах извилины представлена нога, в средних – рука, в нижних – лицо, язык.
Проводники глубокой чувствительности. Ощущение положения и перемещения частей тела в пространстве (кинестезия, мышечно-суставное чувство) базируется на проприоцептивных сигналах. Последние исходят, прежде всего, от рецепторов аппарата движения – нервно-мышечных и нервно-сухожильных веретен, расположенных, соответственно, в мышцах и сухожилиях, связках, суставах. Нервно-мышечные веретена реагируют на растяжение мышцы, нервно-сухожильные
– на ее сокращение. Кроме того, в обеспечении проприоцепции участвуют механорецепторы в капсулах суставов и в связках, рецепторы осязания и давления в коже ладоней, подошв и вокруг суставов, вестибулярные рецепторы. Совокупный анализ всей этой информации позволяет строить представление о схеме тела.
Нервно-мышечное веретено – рецептор, содержащий до 12 специальных поперечнополосатых интрафузальных (внутриверетенных) волокон, в которых нервное волокно разветвляется и образует спиральные и гроздевидные окончания; все эти структуры омываются тканевой жидкостью и заключены в капсулу. Каждая мышца содержит до 50—100 нервно-мышечных веретен, которые фиксированы в ней и растягиваются при удлинении (растяжении) мышцы. При этом спиральные окончания регистрируют степень и скорость растяжения, а гроздевидные – только степень растяжения. Эфферентная иннервация веретен вызывает сокращение последних, тем самым повышая их чувствительность к изменениям длины мышцы. В целом чувствительность этих рецепторов велика. Для мышц плечевого сустава порогом стимуляции является смещение суставного угла на 0,22—0,42° со скоростью 0,3° в секунду; для мышц голеностопного сустава порог смещения равен
1,15—1,3°.
Нервно-сухожильное веретено (сухожильный аппарат Гольджи) – волокна сухожилия, оплетаемые нервными окончаниями и заключенные в капсулу. Эти веретена возбуждаются вследствие сдавливания их пучками сухожильных волокон при натяжении сухожилия, т. е. при сокращении мышцы. В отличие от нервномышечных, нервно-сухожильные веретена не имеют эфферентной иннервации.
Тела первых нейронов пути глубокой чувствительности расположены в спинномозговых ганглиях. Ход волокон глубокой чувствительности до спинного мозга аналогичен прохождению волокон поверхностной чувствительности, но в спинной мозг они вступают не в задние рога, а в задние канатики. Здесь аксоны делятся на короткие и длинные ветви. Короткие ветви, объединяющиеся в ряд пучков (задний продольный пучок Шульца, септомаргинальный пучок Флексига и др.), участвуют в образовании дуги сегментарных (миотатических) рефлексов и формируют межсегментарные связи (не только на своей, но и на противоположной стороне).
Длинные ветви в составе тонкого пучка (Голля) и клиновидного пучка (Бурдаха) в задних канатиках восходят и оканчиваются в продолговатом мозге на телах
вторых нейронов пути глубокой чувствительности, образующих ядра Голля и Бурдаха. Медиально расположенный тонкий пучок проводит импульсы от нижней половины тела (Th5—S5), латерально расположенный клиновидный пучок – от верхней половины (C1—Th4).
Аксоны вторых нейронов пути глубокой чувствительности практически сразу (на уровне олив) переходят на противоположную сторону (образуя медиальную петлю, близко прилегающую к спинномозговой петле) и заканчиваются в таламусе, где в вентролатеральных ядрах расположены тела третьих нейронов пути глубокой чувствительности.
Аксоны третьих нейронов проходят через заднее бедро (заднюю ножку)
внутренней капсулы, лучистый венец и заканчиваются в коре постцентральной извилины. В коре проекция частей тела для глубокой чувствительности такая же, как для поверхностной чувствительности, имеет определенное соматотопическое расположение.
Таким образом, пути проведения поверхностной и глубокой чувствительности имеют идентичное расположение тел первых нейронов (спинномозговые ганглии) и третьих нейронов (таламус), а расположение тел вторых нейронов отличается – собственные ядра задних рогов спинного мозга (поверхностная чувствительность), ядра Голля и Бурдаха (глубокая чувствительность).
Интероцептивная чувствительность в физиологических условиях не осознается. Ощущения, связанные с чувствительностью от внутренних органов (боль, дискомфорт и т. д.), могут возникать при развитии патологического процесса.
Ход проводников мозжечковой рецепции будет представлен в разделе, посвященном мозжечку.
Заключая этот раздел, следует отметить, что большинство представленных выше путей и центров относятся по преимуществу к филогенетически молодой лемнисковой (специфической, информативной) системе. Кроме того, выделяют более древнюю экстралемнисковую (неспецифическую, стимулирующую) систему, которая характеризуется обилием промежуточных инстанций, отдает большое число коллатералей к ретикулярной формации, ее нейроны в таламусе представлены в медиальных ядрах, в коре адресация импульсов менее четкая, импульсы достигают большей площади, информация носит преимущественно качественно-модальный, а не дифференцированный характер.