

Volume1
.pdf48 Часть 1. Введение в мир клетки
возникли как три сравнительно поздние ветви на генеалогическом древе эукариот (см. рис. 1.21). Что касается прокариот, то люди пренебрегали протистами из-за их микроскопических размеров. Только теперь, анализируя геномы, мы начинаем осознавать их положение на древе жизни и оценивать их в контексте нашего отдаленного прошлого согласно тем критериям, которые эти странные существа предлагают нам.
1.3.8. Дрожжи как минимальная модель эукариот
Молекулярная и генетическая сложность эукариот обескураживает. Так что в этом случае еще в большей степени, чем при изучении прокариот, биологам приходится сосредоточивать свои силы и внимание на нескольких отобранных для исследования модельных организмах.
При исследовании внутренней «деятельности» ядерной клетки, чтобы исключить факторы, имеющие отношение к вопросам многоклеточного развития, имеет смысл использовать такой вид, который является не только одноклеточным, но и настолько простым, насколько это возможно. На роль такой минимальной модели эукариот выбрали дрожжевые клетки Saccharomyces cerevisiae (рис. 1.42) — тот же вид, что используется пивоварами для приготовления пива, а пекарями — для выпечки хлеба.
Рис.1.42.ДрожжиSaccharomyces cerevisiae. а)Полученнаянарастровомэлектронноммикроскопефотография скопления клеток. Этот вид клеток известен также как пекарские дрожжи; они размножаются, формируя выпячивание, или почку, которая увеличивается в размере и затем отделяется от материнскойклетки.Намикрофотоснимкевидныпочкующиесяклетки.б)Наполученнойнатрансмиссионном электронном микроскопе фотографии поперечного разреза клетки дрожжей показано ее ядро, мито- хондрияитолстаяклеточнаястенка.(а–любезностьIraHerskowitzиEricSchabatach.)
Итак, S. cerevisiae — это маленький, одноклеточный представитель царства грибов и, согласно современным представлениям, столь же тесно связан с животными, сколь и с растениями; вынослив и легко растет в простой питательной среде. Подобно другим грибам, имеет жесткую клеточную стенку, относительно неподвижен, обладает митохондриями, но не содержит хлоропластов. Когда питательных веществ много, он растет и делится почти так же быстро, как бактерии. Может размножаться вегетативным путем (то есть простым делением клетки) или
Глава 1. Клетки и геномы 49
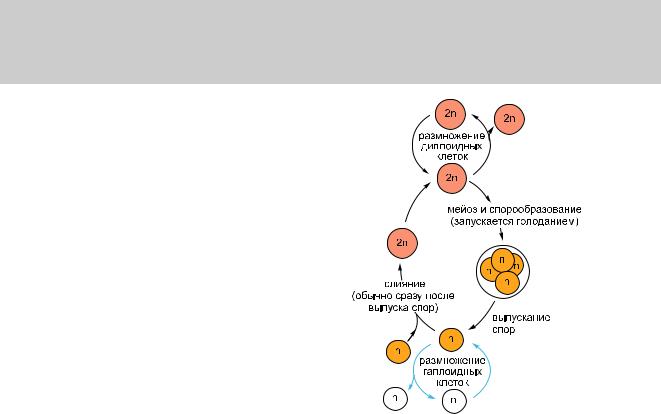
Рис. 1.43. Циклы воспроизводства дрожжей S. cerevisiae. Взависимостиотусловийокружающейсреды и от особенностей генотипа, клетки этого вида могут существовать или в диплоидном (2n) состоянии сдвойнымнаборомхромосом,иливгаплоидном(n) состоянии с одинарным набором хромосом. Диплоидная форма может размножаться обычными циклами деления клетки или предаться мейозу, с тем чтобы произвести гаплоидные клетки. Гаплоидная формаможетилиразмножатьсяобычнымициклами деления клетки, или претерпеть половое слияние с другой гаплоидной клеткой и стать диплоидной. Мейозвызываетсяголоданиемипорождаетспоры— гаплоидные клетки в состоянии покоя, устойчивые кнеблагоприятнымусловиямокружающейсреды.
половым: две гаплоидные (обладают единственной копией генома) клетки дрожжей могут соединиться, чтобы создать клетку, которая будет диплоидной (содержит удвоенный геном); а диплоидная клетка может
подвергнуться мейозу (редукционное де-  ление), чтобы произвести клетки, которые снова окажутся гаплоидом (рис. 1.43). В отличие от высших растений и животных,
ление), чтобы произвести клетки, которые снова окажутся гаплоидом (рис. 1.43). В отличие от высших растений и животных,
дрожжи могут делиться безотносительно своего состояния, переходя то в гаплоидное, то в диплоидное, а процесс, ведущий из одного состояния в другое, может быть вызван изменением условий роста.
В дополнение к этим особенностям дрожжи обладают одним свойством, благодаря которому они стали удобным для генетических исследований организмоммоделью: их геном по эукариотическим меркам очень маленький. Однако он удовлетворяет всем основным требованиям, которым должна соответствовать любая эукариотическая клетка. Как мы увидим позже, исследования, проведенные с использованием дрожжей (S. cerevisiae и других видов), позволили подобрать ключ к открытию многих важнейших клеточных процессов, включая цикл деления эукариотической клетки — это важнейшая цепь событий, в результате которых ядро и все другие компоненты клетки удваиваются и разделяются, чтобы создать две дочерние клетки из одной. Система управления этим процессом была настолько хорошо сохранена в ходе эволюции, что многие из ее компонентов могут функционировать взаимозаменяемо, например, в дрожжах и клетках человека: если у дрожжей-мутантов отсутствует какой-нибудь ген, существенный для цикла деления клетки, то им вводят копию соответствующего гена цикла деления из клетки человека, и дрожжи избавляются от своего «порока» и становятся способными к нормальному делению.
1.3.9. Уровни экспрессии всех генов организма можно отслеживать одновременно
Полная последовательность генома S. cerevisiae, определенная в 1997 г., состоит приблизительно из 13 117 000 пар нуклеотидов, включая маленький вклад

50 Часть 1. Введение в мир клетки
(78 520 п. н.) митохондриальной ДНК. Это общее количество ДНК приблизительно лишь в 2,5 раза больше того, которое имеется у E. coli, а кодирует всего лишь в 1,5 раза больше различных белков (в целом — около 6 300). Образ жизни S. cerevisiae подобен таковому для многих бактерий, и кажется, что эти клетки также испытали на себе давление естественного отбора, что удержало их геном в компактном состоянии.
Знание полной последовательности генома любого организма, будь то дрожжи или человек, открывает новые перспективы для работы с клеткой: то, что когда-то казалось до невозможности сложным, сейчас кажется нам вполне понятным. Теперь есть реальная возможность использовать те методы, которые описаны в главе 8: например, одновременно контролировать количество транскрипта информационной РНК, продуцированной каждым из генов в геноме дрожжей при любых выбранных условиях, и наблюдать, как изменяется эта общая картина активности генов при изменении условий. Анализ можно повторить с информационной РНК, приготовленной из мутантов, не имеющих выбранного нами гена — любого гена, который мы хотим исследовать. В принципе, этот подход позволяет показать полную взаимосвязанную систему управляющих воздействий, которая руководит экспрессией генов не только в клетках дрожжей, но и в любом организме, последовательность генома которого нам известна.
1.3.10. Для постижения устройства клеток нам нужна математика, компьютеры и количественные данные
Пользуясь методами, подобными вышеописанным, и опираясь на нашу информацию о полных последовательностях генома, мы можем составить список генов и белков клетки и приступить к изображению сети взаимодействий между ними (рис. 1.44). Но как мы преобразим всю эту информацию в понятные параметры, описывающие работу клетки? Ведь даже для клеток одного-единственного типа, принадлежащих одному виду организма, современный поток данных кажется непомерным. Неформальная аргументация, на которую обычно опираются в своих рассуждениях биологи, кажется совершенно неадекватной тому объему информации, который надо осмыслить. Фактически дело заключается не только в информационной перегрузке. Биологические системы, например, испещрены контурами обратной связи, и поведение даже наиболее простой из систем с обратной связью чрезвычайно трудно предсказать с помощью одной лишь интуиции (рис. 1.45): небольшие изменения в параметрах могут повлечь за собой коренные изменения всей системы. Для того чтобы перейти от принципиальной схемы, составленной для данной системы, к предсказанию поведения этой системы, нам необходима подробная количественная информация, а для того чтобы на основании полученной информации прийти к адекватным умозаключениям, нам необходимы математика и компьютеры.
Использование таких инструментов для интерпретации результатов на количественном уровне— существенно необходимый, но далеко не всесильный фактор. Может показаться, что, зная, как белки влияют друг на друга и как экспрессия каждого гена регулируется продуктами других генов, нам в скором времени удастся рассчитать, как клетка будет вести себя в целом, подобно тому, как астроном может вычислить орбиты планет, а инженер-химик рассчитать технологические потоки в химической установке. Но любая попытка совершить

Глава 1. Клетки и геномы 51
Рис. 1.44. Сеть взаимодействий между регулирующими гены белками и кодирующими их генами вклеткедрожжей.Результатыпоказаныдля106изобщегоколичества141регулирующихгеныбелков Saccharomycescerevisiae.КаждыйизвключенныхвнаборбелковбылпровереннаспособностьсвязыватьсясрегуляторнойДНКкаждогоизгенов,кодирующихэтотнаборбелков.Насхемегенырасположеныпо кругу,истрелка,направленнаяотгенаАкгенуB,означает,чтобелок,кодируемыйгеномА,связывается с регуляторной ДНК гена B и поэтому, предположительно, регулирует экспрессию гена B. Маленькие кружкисострелкамиуказываютгены,продуктыкоторыхнепосредственнорегулируютихсобственную экспрессию. Гены, управляющие разными моментами поведения клетки, показаны разными цветами. Умногоклеточногорастенияилиживотногочислорегулирующихгеныбелковприблизительнов10раз больше,аколичестворегуляторнойДНК,возможно,в100разбольше,такчтосоответствующаясхемабыла быгораздосложнее.(ИзT. I. Leeetal.,Sciense298:799–804,2002.СразрешенияиздательстваAAAS.)
этот «подвиг» на уровне живой клетки быстро раскрывает ограниченность наших знаний на сегодняшний день. Информация, которой мы обладаем, обильна как таковая, но изобилует пробелами и неясностями. Более того, она в значительной степени носит качественный, а не количественный характер. Наиболее часто цитологи, изучающие системы управления клетки, обобщают свои знания
впростых схематических диаграммах — эта книга полна ими, — а не в числах, графиках и дифференциальных уравнениях. Переход от качественных описаний и интуитивных умозаключений к количественным характеристикам и математической дедукции становится одним из самых больших испытаний для современной цитобиологии. Пока что такое испытание прошло успешно только для нескольких очень простых фрагментов аппарата живых клеток — подсистем, включающих
всебя лишь «горстку» различных белков и два-три взаимно регулируемых гена, где теория и эксперимент могут идти рука об руку. Некоторые из таких примеров мы обсудим в книге позднее.

52 Часть 1. Введение в мир клетки
Рис. 1.45. Довольно простая цепь регулирования гена — единственный ген, регулирующий свою же экспрессию посредством прикрепления своего белкового продукта к своей регу-
ляторной ДНК. Простые схематические представления, такие как это, часто используются, чтобы подытожить все то, что мы знаем (как на рис. 1.44), но оставляют много вопросов безответа.Когдабелоксвязывается,онингибируетилистимулируеттранскрипцию?Насколько
скоростьтранскрипциизависитотконцентрации
белка?Какдолговсреднеммолекулабелкаоста-
етсясвязаннойсДНК?Скольковремениуходит на производство каждой молекулы мРНК или белкаикакбыстродеградируютэтимолекулы?
Математическоемоделированиепоказывает,что,преждечеммысможемпредсказатьповедениедаже такой системы с единственным геном, нам нужно дать «количественные» ответы на все эти и многие другие вопросы. При различных значениях параметров эта система может перейти в уникальное состояние устойчивого равновесия, или может вести себя как переключатель, способный существовать в том или ином наборе альтернативных состояний, или же она может колебаться, или может давать большиеслучайныеколебания.
1.3.11. Из 300 000 видов растений в качестве модели выбран
Arabidopsis thaliana
Большие многоклеточные организмы, которые мы видим вокруг нас: цветы, деревья и животные, — кажутся фантастически разными, но они намного ближе друг к другу по своим эволюционным истокам и более схожи по основам биологии клетки, чем к великому и удивительному хозяину микроскопических одноклеточных организмов. Так, если бактерии и эукариоты отделены друг от друга более чем на 3 000 миллионов лет расходящейся (дивергентной) эволюцией, то позвоночные животные и насекомые отделены друг от друга примерно 700 миллионами лет, рыбы и млекопитающие — примерно 450 миллионами лет, а различные виды цветковых растений — лишь приблизительно 150 миллионами лет.
Ввиду близости эволюционных отношений между всеми цветковыми растениями мы можем еще раз проникнуть в суть клеточной и молекулярной биологии всего этого класса организмов, сосредоточиваясь для детального анализа лишь на одном или нескольких видах. Из нескольких сотен тысяч цветковых растений, населяющих Землю в наши дни, молекулярные биологи сконцентрировали свои усилия на маленьком сорняке, обычной резушке Таля Arabidopsis thaliana (рис. 1.46), который можно выращивать в закрытом помещении в больших количествах и получать тысячи потомков от одного растения через каждые 8–10 недель. Геном арабидопсиса состоит приблизительно из 140 миллионов пар нуклеотидов, что примерно в 11 раз больше, чем у дрожжей, и его полная последовательность уже известна.
1.3.12. Червь, муха, мышь и человек как представители мира животных клеток
Многоклеточные животные составляют большинство всех описанных видов живых организмов, на их изучение приходится бóльшая часть усилий всех прово-

Глава 1. Клетки и геномы 53
Рис.1.46.Arabidopsis thaliana — растение,выбранноевкачестве основной модели для изучения молекулярной генетики расте-
ний.(ЛюбезностьTonyHaydenиJohnInnes.)
димых биологических исследований. В качестве главных модельных организмов для проведения молекулярных генетических исследований выбрано четыре вида. В порядке увеличения размера их располагают в следующий ряд: круглый червь Caenorhabditis elegans, муха Drosophila melanogaster, мышь Mas musculus и человек Homo sapiens. Геномы всех этих видов уже секвенированы.
Caenorhabditis elegans (рис. 1.47) — это ма-
ленький безобидный родственник нематоды, которая повреждает сельскохозяйственные культуры. Представляет собой идеальный модельный организм:
сциклом жизни всего лишь из нескольких дней, со способностью выживать в морозильнике неограниченно долго в состоянии глубокого оцепенения,
спростым строением тела и необычным циклом жизни, — который хорошо подходит для генетических исследований (описаны в главе 23). C. elegans с исправностью часового механизма развивается из оплодотворенной яйцеклетки во взрослого червя в точности с 959 соматическими клетками (плюс изменчивое число яйцеклеток и клеток спермы) — необычная степень регулярности для животного. В настоящее время мы имеем поминутное детальное описание последовательности происходящих событий, таких как деление клеток, их перемещение и изменение характеристик согласно строгим и предсказуемым правилам. Геном из 97 млн. п. н. кодирует приблизительно 19 000 белков, и в распоряжении ученых имеется множество мутантов и других инструментов,
используемых для установления функций различных генов. Хотя общее строение тела червя очень сильно отличается от нашего, сохранение биологических механизмов оказалось для червя достаточным условием, чтобы стать моделью многих процессов, связанных с биологией клетки. Исследования червя помогают нам понять, например, программы деления и некроза клеток, которые определяют число клеток организма, — вопрос большой важности в биологии развития и изучения рака.
1.3.13. Изучение дрозофилы дает ключ к пониманию развития позвоночных
Плодовая мушка Drosophila melanogaster (рис. 1.48) используется в качестве модельного генетического организма дольше, чем какой-либо другой; фактически основы классической генетики были заложены в основном исследованиями этого насекомого. Более 80 лет назад дрозофила обеспечила, например, неопровержимое

54 Часть 1. Введение в мир клетки
Рис.1.47.Caenorhabditiselegans,первыймногоклеточныйорганизм,длякоторогополностьюопреде-
лена последовательность генома. Эта маленькая нематода, около 1 мм в длину, живет в почве. Большинство индивидов — гермафродиты, производящие и яйцеклетки, и сперматозоиды. Приведенное здесь изображение получено с использованием оптического метода интерференционного контраста, благодарячемуяркимицветамичетковырисованыграницытканейживотного;приобычномосвещении животноенеокрашено.(ЛюбезностьIanHope.)
Рис. 1.48. Drosophila melanogaster. Проведенные на этой мухе исследования дали главный ключ к пониманию того, как все животные развиваются из оплодотворенной яйцеклетки во взрослую особь. (ИзE. B. Lewis,Sciense221:cover,1983.СразрешенияиздательстваAAAS.)
доказательство того, что гены — абстрактные единицы наследственной информации — содержатся в хромосомах, конкретных физических объектах, поведение которых точно прослеживалось в эукариотической клетке с помощью светового

Глава 1. Клетки и геномы 55
микроскопа, хотя функция их сначала была неизвестна. Доказательство получили благодаря одной из многих особенностей, которые делают дрозофилу особенно удобным объектом для генетики: в некоторых ее клетках видны гигантские хромосомы
схарактерными полосами (рис. 1.49). Определенные изменения в наследственной информации проявляются в семействах мутантных мух, благодаря чему удалось точно установить корреляцию каждой мутациии с потерей или изменением определенных полос хромосомы-гиганта.
Позже на дрозофиле, больше чем на любом другом организме, было показано, как можно проследить причинно-следственную цепь, ведущую от генетических инструкций, закодированных в хромосомной ДНК, к структуре взрослого многоклеточного организма. Мутанты дрозофилы с расположенными необычным образом частями тела или с неверным рисунком дали ключ для идентификации и характеристики генов, необходимых для создания должным образом структурированного тела:
скишечником, конечностями, глазами и всеми другими частями, расставленными по надлежащим им местам. Как только нуклеотидные последовательности этих генов дрозофилы были определены, появилась возможность провести поиск их гомологов в геномах позвоночных животных. Такие гомологи были найдены, а их функции у позвоночных проверены в ходе анализа мышей, у которых эти гены были видоизменены. Результаты, как мы увидим позднее, показывают удивительную степень подобия молекулярных механизмов развития насекомого и позвоночного.
Большинство всех известных человеку видов живых организмов представлено насекомыми. Так что, даже если бы дрозофила не имела ничего общего с позвоночными животными, а только с насекомыми, она все равно оставалась бы важнейшим модельным организмом. Но если целью является изучение молекулярной генетики позвоночных животных, то почему не заняться этой проблемой прямо? Почему к ней подбираются косвенно, через исследования дрозофилы?
Для развития дрозофилы из оплодотворенного яйца во взрослое насекомое требуется всего лишь 9 дней; размножать ее значительно легче и дешевле, чем любое позвоночное животное, а также ее геном намного меньше — около 170 млн. п. н. по сравнению с 3 200 мил-
Рис.1.49.Гигантскиехромосомыизклетокслюнныхжелез дрозофилы. ПосколькумногораундоврепликацииДНКпрошлобезпромежуточногоделенияклетки,каждаяизхромосомвэтихнеобычныхклеткахсодержитболее1 000идентичных молекул ДНК, выстроенных в цепочку. Благодаря этому
всветовом микроскопе видна характерная для хромосом и воспроизводимая картина полос. Определенные полосы могут быть идентифицированы как местоположения определенных генов: мутантная муха с пропущенной областью
вкартине полос дает представление о фенотипе, обусловленном потерей генов в этой области. Гены, которые транскрибируются с высокой скоростью, соответствуют полосам, которыевыглядят«одутловатыми».Полосы,окрашенныена микрофотографиитемно-коричневымцветом,представляют собойучастки,гдесДНКсвязанспецифическийрегуляторный белок.(ЛюбезностьB. ZinkиR. Paro,изR. Paro,TrendsGenet.6:
416–421,1990.СразрешенияиздательстваElsevier.)

56 Часть 1. Введение в мир клетки
лионами у человека. Ее геном кодирует приблизительно 14 000 белков, и теперь мутанты могут быть получены, по существу, для любого гена. Но есть также и другая, более глубокая причина, по которой генетические механизмы, которые трудно обнаружить у позвоночного животного, часто легко выказываются у мухи. И эта причина кроется, как мы теперь объясняем, в частоте дупликации генов, которая в геномах позвоночных существенно выше, чем в геноме мухи, и, вероятно, была решающим фактором сложного и хитроумного устроения позвоночных.
1.3.14. Геном позвоночных представляет собой продукт повторной дупликации
Почти у каждого гена в геноме позвоночного есть паралоги — другие гены
втом же самом геноме, которые, несомненно, родственны и, должно быть, возникли в результате дупликации генов. Во многих случаях целый кластер генов оказывается близкородственным таким же кластерам, присутствующим в какомлибо другом месте генома, что наводит на мысль о том, что гены дуплицировались не в индивидуальном виде, а связными группами. Согласно одной гипотезе, на раннем этапе эволюции позвоночных полный геном подвергся двум последовательным дупликациям, что положило начало четырем копиям каждого гена. В некоторых группах позвоночных, таких как рыбы семейств лососевых и карповых (в том числе полосатый данио — популярный объект исследований), как предполагают ученые, произошла еще одна дупликация, результатом чего стала восьмикратная представленность генов.
Точный путь эволюции генома позвоночных остается неизвестным, потому что со времени этих стародавних событий произошло много эволюционных изменений. Гены, которые когда-то были идентичными, разошлись; множество копий этих генов было потеряно из-за разрушающих мутаций; некоторые подверглись последующим раундам локальной дупликации; и геном в каждой ветви генеалогического древа позвоночных претерпевал повторные перестройки, почти полностью нарушавшие изначальный порядок генов. Сравнение порядка генов двух эволюционно родственных организмов, таких как человек и мышь, показывает, что часто — во временнóм масштабе эволюции позвоночных — хромосомы сливаются и фрагментируются,
врезультате чего происходит неоднократное перемещение больших блоков последовательности ДНК. Действительно, вполне возможно, как мы покажем в главе 7, что существующее состояние дел — это результат многих отдельных дупликаций фрагментов генома, а не генома целиком.
Однако не вызывает сомнений и тот факт, что дупликация целого генома, на самом деле, время от времени происходит в ходе эволюции, и за такими примерами далеко ходить не надо, так как дублированные наборы хромосом все еще явно опознаются в качестве таковых. Род лягушек Xenopus, например, включает набор близкородственных видов, связанных друг с другом через повторные дупликации или трипликации целого генома. Среди этих лягушек мы можем видеть X. tropicalis с обычным диплоидным геномом; завсегдатая исследовательских лабораторий X. laevis с дублированным геномом и удвоенным количеством ДНК на клетку; и X. ruwenzoriensis с шестикратным дублированием исходного генома и в шесть раз увеличенным количеством ДНК в клетке (108 хромосом по сравнению, например, с 36 у X. laevis). Эти виды, согласно оценкам специалистов, разошлись около 120 миллионов лет назад (рис. 1.50).

Глава 1. Клетки и геномы 57
Рис.1.50.ДвавидалягушкиродаXenopus. X.tropicalis,пред-
ставленныйсверху,имеетобычныйдиплоидныйгеном;X.laevis,помещенныйниже,имеетвдвоебольшееколичествоДНК на клетку. По картинам распределения полос их хромосом и расположению генов на них, а также на основании сравнения последовательностей генов ясно, что вид с бóльшим геномомэволюционировалзасчетдупликациицелогогенома. Этидупликации,какполагают,произошливрезультатеспариваниямеждулягушкамислегкаразошедшихсявидовXenopus. (ЛюбезностьE. Amaya,М. OffieldиR. Grainger,TrendsGenet.14: 253–255,1998.СразрешенияиздательстваElsevier.)
1.3.15. Генетическая избыточность создает лишние проблемы генетикам, но зато дает дополнительные возможности эволюционирующим организмам
Какие бы события ни происходили в эволюционной истории, ясно, что большинство генов в геноме позвоночных существует в нескольких вариантах, когда-то идентичных. Родственные гены зачастую остаются функционально взаимозаменяемыми. Это явление называют генетической избыточностью. Для ученого, изо всех сил пытаю-
щегося обнаружить все гены, участвующие в том или ином процессе, она усложняет задачу. Если ген А мутировал и никакого эффекта не наблюдается, то из этого нельзя заключать, что ген А в функциональном отношении не связан с данным эффектом — просто может оказаться, что этот ген обычно работает параллельно со своими родственниками и их усилий достаточно для практически нормального выполнения функции, даже когда ген А дефектен. В геноме дрозофилы, где дупликация генов происходит реже и повторов в геноме меньше, анализ значительно упрощен: функции отдельных генов проявляются напрямую — по последствиям от мутаций единичных генов (одномоторный самолет падает, когда его двигатель ломается).
Удвоение генома определенно привело к развитию более сложных форм жизни; оно, подобно рогу изобилия, обеспечивает организмы запасными копиями генов, которые могут свободно мутировать и выполнять все более разнящиеся задачи. Пока одна копия оптимизируется, скажем, для использования в печени, другая может оптимизироваться для использования в мозге или приспосабливаться к выполнению новой задачи. Вот почему дополнительные гены усиливают сложность и замысловатость генома в целом. По мере того как гены все больше расходятся
вфункциональном отношении, они перестают быть избыточными. Часто, однако, бывает так, что, несмотря на исполние уже индивидуальных партий, гены продолжают участвовать также и в некоторых эпизодах, выполняя ранее присущую им роль
вмассовке, то есть участвовать избыточно. В таком случае мутация единственного гена вызывает относительно небольшое нарушение, что дает информацию лишь об одной из функций этого гена (рис. 1.51). Семейства генов с разошедшимися, но частично накладывающимися функциями — распространенная особенность мо-