

Volume1
.pdf28 Часть 1. Введение в мир клетки
Рис.1.23.Четыреспособапоявлениягенетических«инноваций»иихвлияниенапоследовательность ДНКорганизма.Когдаклеткидвухразныхтиповвступаютвнерасторжимыйсимбиотическийсоюз,тогда гены одной из них могут переместиться в геном другой — что мы подробно обсудим дальше при рассмотрениимитохондрийихлоропластов.
Дупликация гена: существующий ген может быть продублирован, в результате чего образуется пара близкородственных генов в пределах одной клетки.
Перестройка сегментов: два и более существующих гена могут быть разорваны и воссоединены, так что будет получен гибридный ген, состоящий из сегментов ДНК, которые первоначально принадлежали разным генам.
Горизонтальный (межклеточный) перенос: часть ДНК генома одной клетки может быть передана другой клетке, даже с геномом другого вида. Этот процесс противоположен обычному вертикальному переносу генетической информации, направленному от родителя к потомству.
Изменения всех этих типов оставляют в последовательности ДНК организма характерный след, по которому можно судить о протекании этих четырех процессов. В последующих главах мы обсудим лежащие в их основе механизмы, но пока мы сфокусируем наше внимание на последствиях.

Глава 1. Клетки и геномы 29
1.2.8. Дупликация генов дает начало семействам родственных генов в пределах одной клетки
Каждый раз, когда клетка делится на две дочерние клетки, она полностью удваивает свой геном. Однако иногда происходят «аварии», приводящие к местному дублированию какой-то части генома, при этом в одной клетке остаются и оригинал, и продублированные фрагменты. После подобного удвоения одна из двух копий гена может видоизмениться и начать специализироваться на выполнении особой функции в пределах той же самой клетки. Повторные циклы такого процесса дублирования и расхождения за многие миллионы лет привели к тому, что один ген может дать начало семейству генов, находящихся в пределах одного генома. Анализ последовательности ДНК геномов прокариот показал, что существует множество таких семейств генов: например, у Bacillus subtilis 47 % генов имеет одного или несколько явных родственников (рис. 1.24).
Рис.1.24.СемействаэволюционнородственныхгеноввгеномеBacillussubtilis.Самоепредставительное семейство состоит из 77 генов, кодирующих разновидности ABC-переносчиков — класса мембранных транспортныхбелков,встречающихсявовсехтрехнадцарствахживогомира.(Переработано изстатьи
F. Kunstetal.,Nature390:249–256,1997.СразрешенияMacmillanPublishersLtd.)
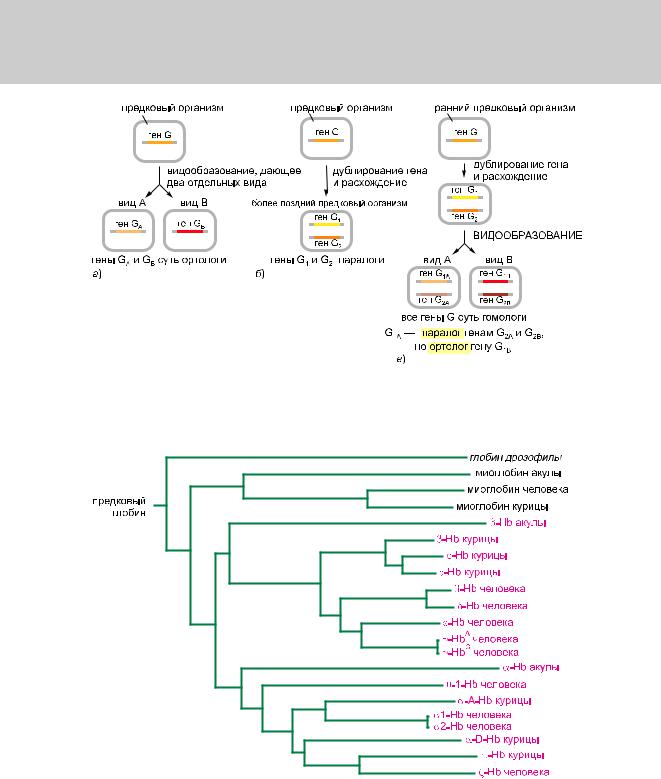
Когда гены подобным образом дублируются и расходятся, особи одного вида наделяются многократными вариантами предкового гена. Этот эволюционный процесс нужно отличать от генетического расхождения, которое происходит, когда один вид разделяется на две отдельные нисходящие линии в точке разветвления в древе семейства: например, когда ветвь человека отделилась от ветви шимпанзе. В ходе эволюции гены постепенно начинают отличаться друг от друга, но, вероятно, продолжают выполнять аналогичные функции в двух сестринских видах. Гены, которые связаны происхождением, то есть гены двух отдельных видов, происходящие от одного и того же гена-предка, который был у последнего общего предка обоих видов, — называют ортологами. Родственные гены, явившиеся результатом дупликации гена в пределах одного генома — и, вероятно, разнящиеся по своим функциям, — называют паралогами. Гены, которые связаны общим происхождением, независимо от пути эволюции, называют гомологами, этот общий термин охватывает связи обоих типов (рис. 1.25).
Взаимоотношения между семействами генов могут оказаться весьма сложными (рис. 1.26). Например, организм, который обладает семейством генов-паралогов
30 Часть 1. Введение в мир клетки
Рис.1.25.Паралогичныеиортологичныегены—дватипагомологиигенов,основаннойнаразличии путейэволюции.а)иб)Основныеварианты.в)Болеесложныйсценарийвозможныхсобытий.
Рис. 1.26. Комплексное семейство гомологичных генов. На этой схеме показана родословная генов гемоглобина(Hb)имиоглобиначеловека,курицы,акулыиглобинадрозофилы.Длиныгоризонтальных линийотражаютстепеньрасхождениямеждуаминокислотнымипоследовательностями.

Глава 1. Клетки и геномы 31
(скажем, 7-ю генами гемоглобина: α, β, γ, δ, ε, ς и θ), может эволюционировать в два отдельных вида (такие как человек и шимпанзе), обладающих полным набором паралогов. Все 14 генов – гомологи, причем ген гемоглобина αчеловека приходится ортологом гемоглобину α шимпанзе, но паралогом гену гемоглобина β человека или шимпанзе, и так далее. Кроме того, гемоглобины позвоночных (белки крови, связыващие кислород) гомологичны миоглобинам позвоночных (мышечные белки, связывающие кислород), а также более отдаленным генам, кодирующим белки, связыващие кислород у беспозвоночных, растений, грибов и бактерий. Обычно по последовательностям ДНК легко распознать гомологию между двумя генами для различных видов; намного труднее, не имея дополнительной информации, решить, состоят ли они в эволюционных отношениях как ортологи.
1.2.9. Гены могут передаваться между организмами как в лаборатории, так и в природе
Прокариоты также дают примеры горизонтальной передачи генов от одних видов клеток к другим. Наиболее явный признак этого – последовательности, распознаваемые по происхождению от бактериальных вирусов, называемых также бак- териофагами (рис. 1.27). Вирусы не являются самостоятельно живущими клетками, но могут выступать в роли векторов для передачи гена: они являют собою маленькие сгустки генетического материала, развившиеся как паразиты на репродуктивных и биосинтетических машинах клеток хозяина. Они копируются в клетке, после этого выходят из нее в защитной оболочке и затем заражают другую клетку, принадлежащую к этому же или другому виду. Часто зараженная клетка уничтожается быстрым и массовым размножением вирусных частиц; но иногда вирусная ДНК, вместо того чтобы непосредственно производить частицы, может сохраняться в организме хозяина на протяжении многих поколений как относительно безвредный пассажир или в виде отдельного внутриклеточного фрагмента ДНК, 

 известного под названием плазмида, или в виде последовательности, вставленной в ненарушенный
известного под названием плазмида, или в виде последовательности, вставленной в ненарушенный
Рис. 1.27. Передача вирусной ДНК от одной клетки другой.
а) Электронная микрофотография частиц вируса бактерий — бактериофагаT4.ГоловкаэтоговирусасодержитвируснуюДНК; хвост содержит аппарат для впрыскивания ДНК в бактериюхозяина. б) Поперечный разрез бактерии, к поверхности которой прикрепился бактериофаг T4. Большие темные пятна внутрибактерии—головкиновыхчастицT4впроцессесборки. Когда они созреют, бактерия разорвется, высвобождая их. (СнимокаперепечатанслюбезногоразрешенияJamesPaulson;
б—JonathanKingиErikaHartwigизG. Karp,CellandMolecular Biology,2nd ed.NewYork:JohnWiley&Sons,1999,исразрешенияиздательстваJohnWiley&Sons.)

32 Часть 1. Введение в мир клетки
геном клетки. В своих путешествиях вирусы могут случайно собрать фрагменты ДНК от генома одной клетки хозяина и переправить их в другую клетку. Такая передача генетического материала часто происходит у прокариот, но также может происходить и между клетками эукариот одного и того же вида.
Горизонтальные переносы генов между клетками эукариот разных видов очень редки и, кажется, не играют существенной роли в их эволюции (хотя, как будет сказано ниже, в эволюции митохондрий и хлоропластов происходили массовые передачи бактериального генома геному эукариот). Напротив, между различными видами прокариот горизонтальный перенос генов происходит намного чаще. Многие прокариоты обладают замечательной способностью подхватывать даже невирусные молекулы ДНК из своего окружения и, таким образом, захватывать генетическую информацию, которую эти молекулы несут. Этим путем или вирус-опосредованным переносом бактерии и археи в природе могут относительно легко приобретать гены соседних клеток. Гены устойчивости к антибиотику или продуцирующие токсин, например, могут быть переданы от одного вида другому и обеспечить преимущество бактерии-получателя в ходе селекции. Этим способом в бактериальных экосистемах эволюционируют новые и иногда опасные штаммы бактерий, которые населяют больницы или различные ниши человеческого организма. Например, горизонтальный перенос генов вызвал распространение в течение последних 40 лет стойких к пенициллину штаммов Neisseria gonorrhoeae — бактерии, которая вызывает гонорею. На более продолжительной шкале времени результаты могут быть еще более впечатляющими: по оценкам ученых, по крайней мере 18 % всех генов современной E. coli приобретено за последние 100 миллионов лет путем горизонтального переноса от других видов.
1.2.10. Половое размножение обеспечивает горизонтальный обмен генетической информацией в пределах вида
Горизонтальный обмен генетической информацией чрезвычайно важен в эволюции бактерий и архей современного мира, и, возможно, в первые дни жизни на Земле этот процесс происходил еще чаще. Существование горизонтального обмена между генами на ранних этапах развития жизни позволило объяснить факты, которые в противном случае озадачили бы любого наблюдателя: эукариоты более близки археям по генам, ответственным за основные процессы обработки информации в ходе репликации ДНК, транскрипции и трансляции, но по генам, ответственным за метаболические процессы, более схожи с бактериями. В любом случае, происходил ли горизонтальный перенос генов свободнее в первые дни жизни на Земле или продолжался с постоянной низкой скоростью на протяжении всей истории эволюции, он здорово усложняет для нас такое понятие, как родословная клетки, так как благодаря ему геном каждой клетки становится соединением частей, полученных из отдельных источников.
Горизонтальный перенос генов среди прокариот может казаться удивительным, но в нем прослеживается аналогия со знакомым нам всем явлением — половым размножением. В дополнение к обычному вертикальному переносу генетического материала — от родителя к потомству — половое воспроизводство становится причиной крупномасштабного горизонтального переноса генетической информации между разными по происхождению клетками, как то: отцовской и материнской. Ключевая особенность полового размножения, конечно, заключается в том, что генетический

Глава 1. Клетки и геномы 33
обмен обычно происходит только между особями одного и того же вида. Но, независимо от того, происходит ли он в пределах вида или между видами, горизонтальный перенос генов оставляет характерный отпечаток: он приводит к появлению особей, которые по одним генам ближе к первой группе родственников, а по другим – ко второй. Сравнивая последовательности ДНК индивидуальных геномов людей, разумный космический пришелец, даже ничего не зная о поведении людей, мог бы сделать заключение, что люди воспроизводятся половым размножением.
Особенно широко половое воспроизводство распространено (хотя и не универсально) среди эукариот. Даже бактерии время от времени позволяют себе управляемый половой обмен ДНК между членами одного и того же вида. Хотя эволюционисты и подвергают сомнению вопрос о преимуществе полового процесса в естественном отборе, но он явно благоприятствовал организмам, способным воспроизводиться половым путем.
1.2.11. Зачастую функция гена может быть установлена по его последовательности
Взаимосвязи генов в пределах семейства важны не только для их исторического значения, но и потому, что они упрощают задачу расшифровки функций гена. После определения последовательности недавно открытого гена ученому достаточно ввести в компьютер всего несколько ключевых последовательностей, для того чтобы в сетевой базе данных провести поиск известных последовательностей генов, родственных «только что обретенному». Во многих случаях функция одного или нескольких таких гомологов уже будет установлена экспериментально, а из этого, так как последовательность гена определяет его функцию, можно предположить функцию нового гена: по всей вероятности, она будет подобна таковой для известных гомологов.
Благодаря этому можно многое разгадать в биологии организма, просто анализируя последовательность ДНК его генома и используя уже известную информацию о функциях генов в других организмах, изученных более детально.
1.2.12. Более 200 семейств генов являются общими для всех трех основных ветвей древа жизни
Располагая полными последовательностями геномов организмов — представителей всех трех надцарств: архей, бактерий и эукариот, — мы можем провести системный анализ гомологичных последовательностей, которые будут перекрывать огромные эволюционные пропасти между ними. Так мы сможем приступить к оценке наследственного материала, общего для всех живых существ. Большие трудности подстерегают исследователя в этом рискованном предприятии. Например, отдельные виды зачастую теряли некоторые предковые гены; другие гены почти наверняка приобретены посредством горизонтального переноса от других видов
ипоэтому не могут считаться истинно предковыми для этих видов, хотя и принадлежат им. Фактически, сравнение геномов настоятельно наводит на мысль, что
иобусловленная происхождением потеря генов, и горизонтальный перенос генов между эволюционно отдаленными видами в каких-то случаях становились главными факторами эволюции, по крайней мере среди прокариот. Наконец, на протяжении 2 или 3 миллиардов лет эволюции некоторые гены, которые были первоначально общими, изменились до неузнаваемости по описанным выше причинам.

34 Часть 1. Введение в мир клетки
Из-за всех этих превратностей эволюционного процесса создается впечатление, что только малая доля предковых семейств генов сохранилась у всех видов в легко узнаваемой форме. Таким образом, из 4 873 кодирующих белок семейств генов, которые идентифицированы при сопоставлении геномов 50 видов бактерий, 13 архей и 3 одноклеточных эукариот, только 63 оказались действительно вездесущими (то есть представленными во всех проанализированных геномах). Значительное большинство этих универсальных семейств относится к компонентам систем транскрипции и трансляции. Вряд ли их можно считать реальным приближением к предковому набору генов. Лучшее — хотя все еще незрелое — представление о последнем может быть получено при регистрации таких семейств генов, которые есть у представителей многих, но не обязательно всех видов из всех трех надцарств. Таким анализом выявлено 264 древних консервативных семейства. Каждому семейству может быть приписана функция (по меньшей мере в плане общей биохимической активности, но, как правило, с большей точностью), при этом наибольшее число генов из этих общих семейств участвует в трансляции, а также в метаболизме и транспорте аминокислот (таблица 1.2). Такой набор высококонсервативных семейств генов представляет собою лишь предварительный набросок наследственного материала, единого для всей современной жизни; дальнейшие успехи в секвенировании геномов и совершенствование методов проведения более тонкого сравнительного анализа, возможно, позволят провести более точную реконструкцию набора генов у последнего общего предка всех живых существ.
1.2.13. Мутации раскрывают функции генов
Не располагая дополнительной информацией, можно сколь угодно долго вглядываться в последовательности геномов, но так и не углядеть функции генов. Конечно, мы можем опознать ген В, подобный гену А, но как распознать функцию гена А изначально? И даже зная функцию гена A, как мы можем проверить, действительно ли функция гена B тождественна ей, как это предполагает подобие их последовательностей? Как совместить мир абстрактной генетической информации с миром реальных живых организмов?
Анализ функций генов зависит от двух взаимодополняющих подходов: генетического и биохимического. Генетика отталкивается от изучения мутантов: мы находим или создаем организм, у которого интересующий нас ген видоизменен,
иисследуем влияние измененного гена на строение и жизнедеятельность организма (рис. 1.28). Биохимия исследует функции молекул: мы извлекаем молекулы из организма и затем изучаем их химические свойства. Объединяя генетику с биохимией
иисследуя химические отклонения в организме мутанта, можно найти именно те молекулы, производство которых зависит от данного гена. В то же время изучение жизнедеятельности организма мутанта показывает нам, какую роль эти молекулы играют в функционировании организма в целом. Таким образом, объединенные усилия генетики и биохимии позволяют связать воедино гены, молекулы, а также строение и функцирование организма.
Впоследние годы информация, полученная о последовательностях ДНК,
имощное совершенствование инструментария молекулярной биологии сделали возможным стремительный прорыв в этой области. На основании сравнительного анализа последовательностей в пределах одного гена часто можно идентифицировать специфические подобласти, которые остались почти неизменными в ходе эволюции. Такие консервативные подобласти, скорее всего, являются самыми важными частя-

Глава 1. Клетки и геномы 35
Таблица1.2.Количествообщихдлявсехтрехнадцарствживогомирасемействгенов,сгруппированных всоответствиисвыполняемымиимифункциями
|
КОЛИЧЕСТВО |
ФУНКЦИЯ СЕМЕЙСТВА ГЕНОВ |
«УНИВЕРСАЛЬНЫХ» |
|
СЕМЕЙСТВ |
Обработка информации |
|
Трансляция |
63 |
Танскрипция |
7 |
Репликация, рекомбинация и репарация |
13 |
Внутриклеточные процессы и передача сигнала |
|
Управление клеточным циклом, митоз и мейоз |
2 |
Защитные механизмы |
3 |
Механизмы передачи сигналов |
1 |
Биогенез между клеточной стенкой и мембраной |
2 |
Внутриклеточный транспорт и выделение |
4 |
Посттрансляционная модификация, белковый обмен, шапе- |
|
роны |
8 |
Метаболизм |
|
Образование и превращение энергии |
19 |
Транспорт и метаболизм углеводов |
16 |
Транспорт и метаболизм аминокислот |
43 |
Транспорт и метаболизм нуклеотидов |
15 |
Транспорт и метаболизм коферментов |
22 |
Транспорт и метаболизм жиров |
9 |
Транспорт и метаболизм неорганических ионов |
8 |
Биосинтез, транспорт и катаболизм вторичных метаболитов |
5 |
Недостаточно охарактеризованные |
|
Предсказаны основные биохимические функции; |
24 |
|
не известна конкретная биологическая роль |
||
|
При проведении данного анализа семейства генов относили к «универсальным», если они были представлены в геномах, по крайней мере, двух различны архей (Archaeoglobus fulgidus и Aeropyrum pernix), двух эволюционно далеких бактерий (Escherichia coli и Bacillus subtilis) и одного эукариота
(дрожжи Saccharomyces cerevisiae). (Данные от R. L. Tatusov, E. V. Koonin и D. J. Lipman, Science 278: 631–637, 1997, с разрешения AAAS; R. L. Tatusov et al., BMC Bioinformatics 4: 41, 2003, с разрешения BioMed Central; и база данных COGs US National Library of Medicine.)
Рис.1.28.Мутантныйфенотип,отражающийфункциюгена.
Нормальные дрожжи (вида Schizosaccharomyces pombe)
по сравнению с мутантами, у которых изменение одного гена привело к преобразованию формы клетки из палочковидной (слева) в Т-образную (справа). Следовательно, мутантныйгенконтролируетформуклетки.Нокакименно науровнемолекулпродуктэтогогенавыполняеттакуюфункцию?Этоболеечемсложныйвопрос,идляответананего необходим биохимический анализ. (Фотография любезно предоставленаKennethSawinиPaulNurse.)

36 Часть 1. Введение в мир клетки
ми гена с точки зрения его функции. Мы можем проверить вклад каждой из них
вактивность продукта гена, вызывая в лабораторных условиях искусственные мутации определенных участков в пределах гена или конструируя искусственные гибридные гены, в которых сочетаются части разных генов. Дабы облегчить биохимический анализ, можно конструировать организмы, в больших количествах синтезирующие как РНК, так и белки, предписываемые этим геном. Специалисты
вобласти структуры молекул могут определить трехмерную конформацию продукта гена, показывающую точное положение каждого атома в нем. Биохимики могут определить, какой вклад каждая из частей генетически предписанной молекулы вносит в ее химическое поведение. Цитологи могут анализировать поведение клеток, сконструированных для экспрессии мутантного гена.
Однако нет ни простого рецепта открытия функции гена, ни простой стандартной универсальной формы для ее описания. Например, мы можем обнаружить, что продукт данного гена катализирует некоторую химическую реакцию, но при этом не иметь ни малейшего представления о том, как или почему данная реакция важна для организма. Функциональная характеристика каждого нового семейства продуктов гена, в отличие от описания последовательностей генов, представляет собой новоявленное испытание находчивости биолога. Кроме того, мы никогда полностью не постигнем функции гена до тех пор, пока не изучим его роль в жизни организма как единого целого. Для того чтобы получить полное представление о функциях гена, нам следует изучать организмы в целом, а не только молекулы или клетки.
1.2.14. Молекулярные биологи сфокусировали свое внимание на Escherichia coli
Живые организмы настолько сложны, что чем больше мы узнаем о каких-либо отдельных видах, тем более привлекательными они становятся в качестве объекта дальнейшего изучения. Каждое открытие поднимает новые вопросы и предоставляет новый инструментарий для работы над общими вопросами в контексте выбранного организма. По этой причине большие коллективы биологов посвящают свою жизнь изучению одного и того же модельного организма во всех его проявлениях.
Вчрезвычайно многообразном мире бактерий внимание молекулярных биологов
втечение долгого времени концентрировалось только на одном виде: Escherichia coli, или E. coli (см. рис. 1.17 и 1.18). Эта маленькая, имеющая форму прута одноклеточная бактерия обычно живет в кишечнике человека и других позвоночных животных, но может быть и легко выращена — в простом питательном бульоне
вкультуральном сосуде. Она приспосабливается к изменяющимся химическим условиям, быстро размножается и может эволюционировать посредством мутаций и отбора с поразительной скоростью. Как и у других бактерий, различные штаммы E. coli, хотя их и классифицируют как представителей одного вида, отличаются генетически в большей степени, чем различные разновидности воспроизводящегося половым путем организма типа растения или животного. Один штамм E. coli может обладать многими сотнями генов, которые отсутствуют у другого, а у двух штаммов может быть не более 50 % общих генов. Стандартный искусственный штамм E. coli K-12 имеет геном приблизительно из 4,6 млн. п. н., содержащихся в единственной кольцевой молекуле ДНК, кодирующей приблизительно 4 300 различных видов белков (рис. 1.29).
На молекулярном уровне мы знаем об E. coli больше, чем о любом другом живом организме. Бóльшая часть из постигнутого нами о фундаментальных ме-

Глава 1. Клетки и геномы 37
Рис.1.29.ГеномE. coli. а)СкоплениеклетокE.coli.б)СхемагеномаштаммаK-12E.coli.Схемаявляется круговой,потомучтоДНКE.coli,подобнотаковойостальныхпрокариот,образуетединичнуюзамкнутую петлю.Кодирующиебелокгеныпоказаныжелтымиилиоранжевымиполоскамивзависимостиотнити ДНК,скоторойонитранскрибируются;гены,кодирующиетолькомолекулыРНК,обозначенызелеными стрелками.ОднигенытранскрибируютсясоднойцепидвойнойспиралиДНК(внаправлениичасовой стрелкинаэтойсхеме),другие—сдругойнити(противчасовойстрелки).(Снимокалюбезнопредоставлен докторомTonyBrainиDavidParker(фотолаборанты);изображениебпереработаноизстатьиF. R. Blattner etal.,Science277:1453–1462,1997.СразрешенияиздательстваAAAS.)
ханизмах жизни, таких как реплицирование клетками содержащейся в них ДНК или расшифровка инструкций, заложенных в ДНК, для направленного синтеза определенных белков, получена в ходе изучения E. coli. Основные генетические механизмы оказались сильно консервативными на протяжении всей эволюции, поэтому в наших клетках эти механизмы по существу те же, что и у E. coli.
Заключение
Прокариоты (клетки без обособленного ядра) — это самые разнообразные в биохимическом отношении организмы и включают виды, которые могут по- лучать всю свою энергию и питательные вещества из неорганических химиче- ских источников, таких как реакционно-способные смеси минеральных веществ, выбрасываемые гидротермальными источниками океанического ложа, — такой «диеты», возможно, придерживались первые живые клетки 3,5 миллиарда лет
тому назад. Сравнительный анализ последовательностей ДНК выявляет «се- мейные отношения» между живыми организмами и показывает, что прокариоты подразделяются на две группы, которые образовались на заре эволюции: бактерии (или эубактерии) и археи. Вместе с эукариотами (клетки с окруженным мем- браной ядром) они составляют три первичные ветви древа жизни. Большинство бактерий и архей — это маленькие одноклеточные организмы с компактными