

1 курс / Гистология / Miadelets-OD_Gistologiia_tsitologiia_i_embriologiia_cheloveka_Ch-2_2016
.pdfхелперам. МНС II класса взаимодействуют с Т-клеточным рецептором и CD4+ на поверхности Т-хелперов CD4+.
Всю паракортикальную зону можно разделить на отдельные единицы. Каждая единица прилежит к лимфоидному узелку коркового вещества и состоит из центральной и периферической частей. В центральной части происходит бласттрансформация и размножение Т-лимфоцитов. На периферии находятся посткапиллярные вены (ПКВ) с высоким эндотелием. Через них происходит миграция лимфоцитов из крови в лимфоузел и, по мнению некоторых авторов, в обратном направлении. Вместе с тем, часть лимфоцитов, в первую очередь Т-клетки, поступают в лимфоузел по приносящим лимфатическим сосудам, а не из крови через ПКВ.
ПКВ с высоким эндотелием имеют общий диаметр от 12 до 20 мкм и диаметр просвета от 5 до 9 мкм. Эндотелиоциты ПКВ характеризуются рядом специфических морфофункциональных свойств. Их цитоплазма имеет умеренную электронную плотность и содержит значительное количество органелл, концентрированных в околоядерной части. Однако и в других участках цитоплазмы их содержание достаточно высокое. Митохондрии располагаются равномерно. Достаточно хорошо развит комплекс Гольджи. Лизосомы единичны. Гранулярная ЭПС развита гораздо лучше, чем в эндотелиоцитах другой локализации. Между эндотелиоцитами имеются плотные контакты и десмосомы. В эндотелиоцитах содержится значительное количество микропиноцитозных везикул, а также телец Паладе-Вейбеля (см. ниже). Фенестры в эндотелии единичны. Ядра клеток имеют неправильную, часто лопастную форму, с ядрышками и крупнозернистым гетерохроматином. Эндотелий расположен на выраженной базальной мембране, не имеющей пор и истончений.
ПВК паракортикальной зоны способны к активации, которая сопровождается появлением на их цитолемме различных молекул клеточной адгезии. Один из механизмов активации эндотелиоцитов опосредуется синтезируемыми локально цитокинами, такими, как интерферон , ИЛ-1 и фактор некроза опухолей (ФНО).
Активированные эндотелиоциты ПКВ из плоских превращаются в кубические и начинают экспрессировать ряд молекул межклеточной адгезии суперсемейства иммуноглобулинов. Экспрессируются также адгезионные молекулы семейства селектинов, в том числе Е-селектин ELAM-1 (CD62E) и Р-селектин (СD62P). Р-селектин хранится в особых органеллах эндотелиоцитов - тельцах Паладе-Вейбеля - и при активации эндотелия быстро транспортируется на его поверхность.
Ведущая роль в адгезии лейкоцитов к эндотелиоцитам отводится CD44, который экспрессируют все лейкоциты. Как полагают, между лимфоцитами и эндотелиоцитами возникают адгезионные взаимодействия, в результате которых лимфоциты направляются в соответствующие ткани-мишени, в
371
данном случае в ткани лимфоузла. Это происходит за счет экспрессии эндотелием специфических для лимфоузла “адрессинов”.
Высота эндотелиоцитов может изменяться, что влияет на интенсивность миграционных процессов: при высоком эндотелии между эндотелиоцитами появляются широкие щели, через которые осуществляется максимальная миграция клеток. При снижении высоты эндотелиоцитов межклеточные пространства исчезают, что ведет к свертыванию миграционных процессов.
ПОСЛЕДОВАТЕЛЬНОСТЬ ИММУННЫХ РЕАКЦИЙ В ПАРАКОРТИКАЛЬНОЙ ЗОНЕ ПРИ КЛЕТОЧНОМ ИММУНИТЕТЕ.
Антигены поступают в паракортикальную зону лимфоузлов с током лимфы по лимфатическим синусам. В качестве антигенов могут выступать либо отдельные клетки (например, клетки трансплантата), либо слущившиеся с поверхности трансформированных или чужеродных клеток молекулы. Кроме того, антиген транспортируется в лимфоузел дендритными клетками Лангерганса.
Впаракортикальной зоне происходит антигензависимая дифференцировка Т-лимфоцитов после попадания в эту зону антигена. Микроокружением для лимфоцитов здесь являются свободные макрофаги и интердигитирующие клетки (ИДК). Одновременно с антигенами MHC I класса ИДК экспрессируют и белки MHC II класса. Эти клетки обрабатывают антиген до высокоиммунной формы и в комплексе с молекулами МНС I класса выделяют на свою поверхность, одновременно секретируя ИЛ-1. Эти два стимула активируют Т-хелперы (Тх1), которые, выделяя ИЛ-2, вызывают бласттрансформацию Т-лимфоцитов. После размножения последние дифференцируются в цитотоксические Т-лимфоциты (киллеры), Т-хелперы и Т- клетки памяти.
Впоследнее время твердо установлено, что реакции клеточного иммунитета протекают не только с участием клеточных факторов иммунитета (Т- лимфоциты, NK-клетки, макрофаги, гранулоциты), но и гуморальных факторов, в частности, антител и многочисленных цитокинов (например, антителозависимая цитотоксичность). Показано, что в реакциях клеточного им-
мунитета участвуют не только цитотоксические (киллерные) Т-лимфоциты, несущие маркер СD8+ и распознающие антиген в комплексе с молекулами MHC I класса, но и хелперные СD4+ лимфоциты, распознающие антиген в комплексе с молекулами МНС II класса и осуществляющие лизис чужеродных или измененных собственных клеток через включение в них рецепторно опосредованного апоптоза. Обе разновидности Т-лимфоцитов образуются в паракортикальной зоне лимфоузлов. Как отмечалось, цитолиз могут осуществлять также макрофаги, НК-, К-клетки и гранулоциты.
К морфологическим проявлениям иммунного ответа по клеточному типу относится увеличение объема паракортикальной зоны, появление большого количества Т-лимфобластов и делящихся лимфоцитов. Может наблю-
372
даться ситуация, когда паракортикальная зона увеличивается особенно резко. Это приводит к трансформации и коркового вещества, заключающейся в том, что лимфоидные узелки сливаются в монолитное плато. При удалении тимуса эта зона, напротив, атрофируется.
МОЗГОВОЕ ВЕЩЕСТВО состоит из двух структурно-функциональных компонентов: мозговых (мякотных) тяжей и мозговых промежуточных синусов. Мозговые тяжи являются В-зависимой зоной. Здесь происходит окончательное созревание мигрировавших из коры предшественников плазмоцитов в плазмоциты. Накапливающиеся при иммунном ответе в мозговых тяжах плазмоциты секретируют в лимфу антитела. Снаружи к мозговым тяжам прилежат мозговые синусы.
СТРОЕНИЕ СИНУСОВ ЛИМФОУЗЛА. Все синусы лимфоузла представляют щелевидные пространства, которые выстланы эндотелием, способным к фагоцитозу (эндотелиоциты синусов лимфоузла часто называют
“береговыми”, литоральными клетками). Некоторые авторы считают
“береговые” клетки ретикулярными клетками. Кроме эндотелиоцитов, в образовании стенки лимфатических синусов узла участвуют рететелиальные клетки. Эти клетки имеют отростчатую форму, причем отростки, направленные в противоположные стороны, пересекают пространство синуса и, подойдя к его стенкам, формируют расширения в виде площадок, которые наряду с литоральными клетками участвуют в образовании прерывистой выстилки синусов (в субкапсулярном синусе со стороны капсулы выстилка непрерывна). Базальная мембрана в выстилке синусов отсутствует. Отростки рететелиальных клеток формируют в просвете синуса трехмерную сеть, замедляющую ток лимфы, что способствует ее более полному очищению макрофагами. Идущие в разных направлениях ретикулярные волокна также формируют сеть. Эти две сети замедляют ток лимфы и (особенно) клеток по синусам и способствуют ее санации, а также развертыванию иммунного ответа. В синусах содержится большое количество свободных макрофагов и лимфоцитов, которые могут фиксироваться на сети.
КРОВОСНАБЖЕНИЕ ЛИМФАТИЧЕСКОГО УЗЛА. Кровеносные сосуды входят в ворота узла. От артерий отходят капилляры в капсулу и трабекулы, а также к узелкам. Вокруг узелков и в них формируются поверхностная и глубокая капиллярная сети. Капиллярные сосуды продолжаются в венулы с высоким эндотелием, а затем в вены, которые выходят через ворота узла. В норме кровь никогда не поступает в синусы. При воспалении, травмах и других патологических состояниях подобное явление возможно.
ИННЕРВАЦИЯ ЛИМФАТИЧЕСКОГО УЗЛА. Афферентная иннервация лимфоузла обеспечивается клетками Догеля II типа и псевдоуниполярными нейронами соответствующих спинальных ганглиев. Эфферентная иннервация включает симпатическое и парасимпатическое звенья. Имеются мелкие интрамуральные ганглии. Нервы входят в лимфоузел по ходу сосудов, в ад-
373
вентиции которых формируют обильную сеть. От этой сети отходят веточки, направляющиеся в мозговое и корковое вещество. В последнем они лежат в основном в межузелковой части либо концентрически по периферии узелков. Внутри узелков нервные структуры отсутствуют. Очень часто с холинергическими и адренергическими нервными волокнами контактируют крупные клетки типа макрофагов и тканевых базофилов. В последнее время существуют представления о так называемых “открытых синапсах”, когда нервное окончание, подойдя к группе иммунокомпетентных клеток, выделяет медиатор, диффундирующий между клетками и оказывающий на них регулирующее влияние. Стимуляция симпатического звена ВНС приводит к некоторому угнетению иммунного ответа в результате накопления в иммунокомпетентных клетках циклического АМФ, тормозящего бласттрансформацию и деление лимфоцитов. Стимуляция парасимпатического звена вызывает противоположный эффект.
РЕГЕНЕРАЦИЯ. Физиологическая регенерация лимфоузлов протекает на высоком уровне. В ее основе лежит деление и дифференцировка лимфоцитов и ретикулярных клеток. Посттравматическая регенерация может происходить лишь в том случае, если сохраняются приносящие и выносящие лимфатические сосуды с прилежащей к узлу соединительной тканью. В основе посттравматической регенерации, так же, как и физиологической, лежит пролиферация вначале клеток ретикулярной ткани, а затем и лимфоцитов.
ВОЗРАСТНЫЕ ИЗМЕНЕНИЯ. К моменту рождения лимфатические узлы не достигают окончательного развития, их окончательное структурное созревание происходит в раннем постнатальном периоде. Лимфоузлы новорожденного полиморфны, богаты лимфоцитами. Синусы часто не определяются из-за переполнения лимфоцитами. Фолликулы с центрами размножения редки. Плазматические клетки обычно не выявляются. Могут встречаться клетки миелоидного кроветворения. В течение первого года жизни число вторичных лимфоидных узелков нарастает. Появляются и быстро увеличиваются в количестве плазматические клетки. Новообразование вторичных узелков особенно активно в возрасте 4-8-и лет, а к 12-и годам дифференцировка лимфоузлов в основном заканчивается. При старении вновь исчезают лимфоидные узелки с центрами размножения. Одновременно утолщается капсула лимфоузлов. Уменьшается количество макрофагов и их фагоцитарная активность. Некоторые узлы почти полностью атрофируются и замещаются жировой тканью. Вместе с тем, во многих лимфоузлах их клеточный состав сохраняется даже у глубоких стариков.
КЛИНИЧЕСКОЕ ЗНАЧЕНИЕ ЛИМФОУЗЛОВ. Лимфатические узлы, находясь по току лимфы, очищают ее от всех патогенных агентов. Поэтому они являются непосредственными участниками иммунных реакций, являясь вторым барьером на пути антигенов (первым барьером являются кожа и сли-
374
зистые оболочки внутренних органов). Вместе с тем, в самих лимфоузлах при массивной антигенной атаке могут возникать очаги инфекции. В лимфоузле могут также задерживаться, а затем и пролиферировать метастазирующие клетки злокачественных опухолей. Поэтому при радикальных оперативных вмешательствах по поводу злокачественных опухолей хирурги стремятся удалить регионарные лимфоузлы на тот случай, если в них уже мигрировали опухолевые клетки. Системное увеличение лимфатических узлов наблюдается при опухолях лимфоидной ткани, например, при лимфо-
грануломатозе, лимфолейкозе и др.
ГЕМОЛИМФАТИЧЕСКИЕ УЗЛЫ. У некоторых млекопитающих (жвачные, свинья и др.), а также у человека (редко) встречаются гемолимфатические узлы. Гемолимфатическими называются узлы, которые связаны не только с лимфатическими, но и с кровеносными сосудами. В них отсутствует четкое разделение на кору и мозговое вещество. Гемолимфатические узлы чаще располагаются в жировой клетчатке по ходу почечных артерий, аорты, иногда в заднем средостении. Снаружи они покрыты соединительнотканной капсулой. Корковое вещество меньше по объему, содержит меньшее, чем лимфатические узлы, количество узелков. Менее выражены и малочисленны мякотные тяжи мозгового вещества. Синусы, особенно мозговые, напротив, хорошо выражены, содержат кровь. Их трудно отличить от переполненных кровью вен. В качестве критерия служат находящиеся в просвете синусов ретикулярные клетки и ретикулярные волокна. В гемолимфатических узлах, так же, как и в лимфатических, имеются приносящие и выносящие сосуды.
В гемолимфатических узлах наряду с лимфопоэзом, который преобладает, встречаются клетки миелоидного ряда: гранулопоэтического, эритропоэтического и тромбоцитопоэтического рядов. У жвачных животных гемолимфатическе узлы являются добавочными селезенками, т.е. включены не в лимфатическое, а в кровеносное русло. Поэтому в них отсутствуют приносящие и выносящие лимфатические сосуды. Кроме того, в добавочных селезенках лимфоидные узелки имеют строение, свойственное для таковых в се-
лезенке: в них есть центральные артерии и периартериальные лимфоидные муфты (ПАЛМ), артериальные гильзы, венозные синусы (см.
строение селезенки), которые отсутствуют в истинных гемолимфатических узлах. Истинные гемолимфатические узлы следует отличать от ложных, образующихся в результате всасывания крови лимфатическими сосудами и поступления ее в лимфоузлы из очагов кровоизлияний. В таких лимфоузлах в отличие от истинных гемолимфатических узлов кровь обнаруживается в краевом синусе и в афферентных лимфатических сосудах.
С возрастом гемолимфатические узлы в гораздо более выраженной степени, чем лимфатические узлы, подвергаются инволюции. Их корковое и мозговое вещество замещается жировой тканью или РВНСТ.
375
СЕЛЕЗЕНКА
ФУНКЦИИ СЕЛЕЗЕНКИ.
1.Кроветворная - образование лимфоцитов в ходе антигензависимого лимфопоэза.
2.Барьерно-защитная - осуществление неспецифических защитных реакций (фагоцитоз) и специфических иммунных реакций. Селезенка находится по току крови и очищает ее от антигенов за счет деятельности многочисленных макрофагов. Наряду с этим, в селезенке синтезируются необходимые для уничтожения антигенов антитела. Селезенка участвует и в реакциях клеточного иммунитета. В последнее время при сепсисе, когда в силу перегруженности макрофагальной системы селезенки элиминация из кровотока бактерий осуществляется неэффективно и они беспрепятственно размножаются в крови, с лечебной целью применяют сеансы подключения в сосудистую систему человека нативной селезенки свиньи. Это лечебное мероприятие оказывается настолько эффективным, что больные полностью выздоравливают после нескольких сеансов. Данный пример показывает ту роль, которую играет селезенка в сохранении антигенного гомеостаза организма. Поэтому в последнее время хирурги стараются как можно реже и только по строгим показаниям прибегать к удалению селезенки (спленэк-
томии).
3.Депонирующая функция. Селезенка является депо крови. В состоянии покоя организма в селезенке может депонироваться определенное количество крови, которая при переходе к состоянию активного бодрствования выбрасывается в общий кровоток. При наложении лигатуры на селезеночную артерию масса селезенки сокращается в 4 раза. Кроме крови, в селезенке депонируются и концентрируются тромбоциты (до 1/3 от их общего количества в организме).
4.Обменная функция. Селезенка регулирует обмен углеводов, железа, стимулирует синтез белков, факторов свертывания крови, холестерина и билирубина, влияет на синтетические функции печени.
5.Гемолитическая функция. При участии лизолецитина селезенка разрушает старые эритроциты. Поэтому иногда селезенку называют кладбищем эритроцитов. Высвобождающееся при этом железо связывается с транспортным белком трансферрином и транспортируется в костный мозг, где включается в гемоглобин вновь образующихся эритроцитов. Одновременно в селезенке разрушаются стареющие и поврежденные тромбоциты. При патологии в селезенке может происходить массовая гибель нормальных тромбоцитов, что ведет к повышенной кровоточивости и анемии. В таких случаях иногда прибегают к удалению селезенки. Это ведет в восстановлению числа тромбоцитов и даже к тромбоцитозу - увеличению их числа выше нормы.
376

6. Эндокринная функция - синтез эритропоэтина, стимулирующего эритропоэз. Кроме того, в селезенке вырабатываются вещества тафтсин и спленин. Тафтсин - пептид, стимулирующий активность фагоцитов. При удалении селезенки содержание тафтсина резко падает, что ведет к снижению резистентности организма к инфекциям. Спленин является аналогом тимопоэтина. Он стимулирует бласттрансформацию и дифференцировку Т- лимфоцитов.
РАЗВИТИЕ. Закладка селезенки происходит на 5-й неделе эмбриогенеза. В толще мезенхимы дорзальной брыжейки образуется плотное скопление мезенхимы, которая превращается в ретикулярную ткань, прорастает кровеносными сосудами и заселяется стволовыми кроветворными элементами. На 5-м месяце эмбриогенеза в селезенке происходит и миело-, и лимфопоэз, затем миелопоэз постепенно прекращается, и селезенка становится органом лимфопоэза. У некоторых лабораторных животных (например, мыши) миелопоэз сохраняется и во взрослом состоянии. Возобновление в селезенке человека миелопоэза может происходить при патологии.
СТРОЕНИЕ (Рис. 21.11). Селезенка является паренхиматозным зональным органом. Снаружи она покрыта соединительнотканной капсулой, к которой прилежит мезотелий. Капсула содержит гладкие миоциты. От капсулы отходят трабекулы из РВНСТ. В трабекулах, как и в капсуле, встречаются пучки гладких миоцитов. Капсула и трабекулы образуют опорносократительный аппарат селезенки и составляют до 7% ее объема. Все пространство между капсулой и трабекулами заполнено ретикулярной тканью. Ретикулярная ткань, трабекулы и капсула образуют строму селезенки.
Совокупность лимфоидных клеток представляют ее паренхиму.
Рис. 21.11. Строение селезенки.
А-В – фрагменты органа; 1 – мезотелий се-
розной оболочки; 2
– капсула; 3 – трабекулы; 4 - лимфоидный узелок (компонент белой пульпы) и в нем: 5 – центр размножения; 6 – мантийная зона; 7 – маргинальная зона белой
пульпы; 8 – периартериолярная лимфоидная муфта; 9 - центральная артериола; 10 – синусоид селезенки; 11 - селезеночный тяж; 12 - трабекулярные артерия и вена
Вселезенке выделяют две различающиеся по строению зоны - крас-
ную и белую пульпу.
Белая пульпа представляет собой совокупность лимфоидной ткани в виде узелков, лежащих вокруг центральных артерий. Ее общий объем равен 1/5 части селезенки. Лимфоидные узелки селезенки отличаются по строению от узелков лимфоузла, т.к. содержат и Т-, и В-зоны (напомним, что в лимфоузле фолликул является В-зоной). Каждый лимфоидный узелок имеет 4 зоны:
1. Реактивный центр (центр размножения).
2. Мантийная зона - корона из малых в-лимфоцитов памяти. 3. Маргинальная зона.
4. Периартериальная зона, или периартериальная лимфоидная муфта (ПАЛМ) - зона вокруг центральных артерий.
1-я и 2-я зоны соответствуют лимфоидным узелкам лимфоузла и являются
В-зоной селезенки.
Вцентре размножения лимфоидных узелков происходит бласттрансформация и пролиферация В-лимфоцитов. Здесь находятся антигенпредставляющие фолликулярные дендритные клетки, В-лимфоциты на разных стадиях развития и делящиеся В-лимфоциты, претерпевшие бласттрансформацию. В мантийной зоне происходит кооперация Т- и В-лимфоцитов и накопление В-лимфоцитов памяти.
Т-лимфоциты, составляющие около 60% всех лимфоцитов белой пульпы, лежат вокруг центральной артерии в 4-й зоне, поэтому данная зона является Т-зоной селезенки. При клеточном иммунном ответе эта зона подвергается гипертрофии. В зоне ПАЛМ находятся интердигитирующие клетки и свободные макрофаги, осуществляющие процессинг антигенов притекающей крови и презентирующие их в комплексе с молекулами МНС I класса и ИЛ-1 Т-хелперам, запуская реакции клеточного иммунитета.
Снаружи от ПАЛМ и мантийной зон узелков находится маргинальная зона. Ее окружает маргинальный синус. В этой зоне происходят кооперативные взаимодействия Т- и В-лимфоцитов, через нее из крови в белую пульпу поступают Т- и В-лимфоциты, заселяющие впоследствии Т- и В- зоны, а также антигены, которые здесь захватываются макрофагами. Через эту зону в красную пульпу мигрируют созревающие плазмоциты. Клеточный состав маргинальной зоны представлен лимфоцитами (большей частью В- лимфоциты и предшественники плазмоцитов), макрофагами, ретикулярными клетками.
Красная пульпа селезенки состоит из:
Пульпарных синусов;
378
Пульпарных (бильротовых) тяжей.
Пульпарные тяжи в своей основе содержат ретикулярную ткань. Между ретикулярными клетками находятся эритроциты (часть из них в состоянии распада), зернистые и незернистые лейкоциты, плазмоциты на разных стадиях созревания. Функциями пульпарных тяжей являются:
1)распад и уничтожение старых эритроцитов;
2)созревание плазмоцитов (в определенной степени пульпарные тяжи аналогичны мозговым тяжам лимфоузла);
3)осуществление обменных процессов.
В красной пульпе находится множество макрофагов, которые очищают кровь от содержащихся в ней антигенов.
Синусы красной пульпы - это часть кровеносной системы селезенки. Они составляют большую часть красной пульпы. Синусы имеют диаметр 1240 мкм и относятся к венозной системе, однако по строению близки к синусоидным капиллярам: выстланы эндотелием, который лежит на прерывистой базальной мембране. Эндотелиоциты имеют веретеновидную или палочковидную форму, лежат продольно, неплотно друг к другу (имеются щели) и окружены циркулярными ретикулярными волокнами, в результате чего формируют картину “рассохшейся деревянной бочки”. Кровь из синуса может прямо поступать в ретикулярную основу селезенки. Функции синусов - проведение крови, ее депонирование, обеспечение обмена кровью между сосудистой системой и стромой красной пульпы.
Соотношение белой и красной пульпы может быть различно. В связи с этим выделяют два типа селезенок:
а) иммунный тип характеризуется выраженным развитием белой пуль-
пы;
б) метаболический тип, при котором значительно преобладает красная пульпа, осуществляющая обменные процессы.
КРОВООБРАЩЕНИЕ В СЕЛЕЗЕНКЕ (Рис. 21.12). Кровь поступает в селезенку по селезеночной артерии, которая распадается на сегментарные,
трабекулярные и пульпарные артерии. Трабекулярные артерии лежат в трабекулах, пульпарные - в красной пульпе селезенки. Пульпарная артерия, которая входит в белую пульпу, называется центральной артерией. Она является артерией мышечного типа. В центре узелка она располагается только в эмбриональном периоде, когда вокруг нее сконцентрировано
379
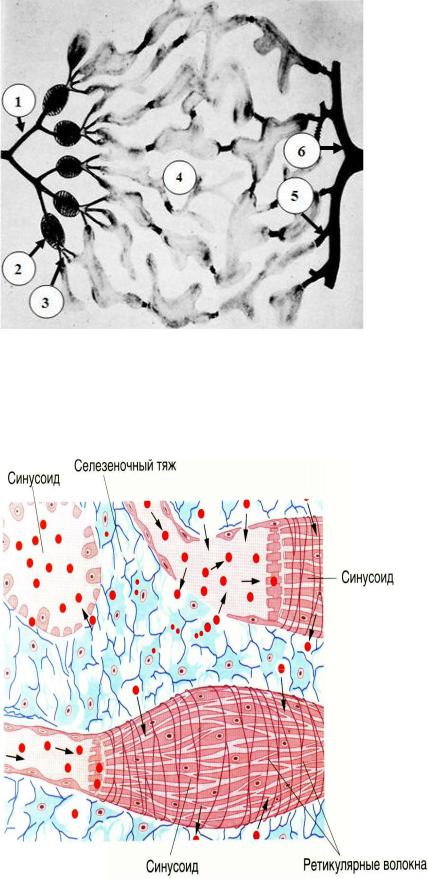
Рис. 21. 12. Схема кровоснабжения селезенки 1 – пульпарная артерия; 2 –
лимфоидный узелок; 3 – кисточковые артериолы; 4 – пульпарные синусы; 5 – пульпарная вена; 6 – трабекулярная вена
небольшое количество лимфоцитов. После рождения артерия оттесняется на периферию заселившимися Т- лимфоцитами. В белой пульпе от центральной артерии отходят капилляры, снаб-
жающие кровью белую пульпу, расходящиеся радиально и проникающие вплоть до маргинальной зоны узелка, где вливаются в маргинальный синус. Основной ствол центральной артерии, выходя из узелка в красную пульпу (или прямо в нем), распадается на несколько артериол, которые формируют подобие кисти руки и называются кисточковыми. Артериолы ок-
ружены периарте-
риолярными макрофагальными муфтами (эллипсои-
дами). В этом участке в стенке артериолы отсутствуют миоциты, базальная мембрана эндотелия прерывистая, а стенку эллипсоида образуют макрофаги с высокой фагоцитарной активностью. В состав эллипсоидов входят также лимфоциты. Считается, что эллипсоиды участвуют в фильтрации и де-
зинтоксикации крови, протекающей через селезенку. Кисточковые артериолы продолжаются в венозные синусы красной пульпы (Рис. 21.13).
380