

Проблема митогенетического излучения как аспект молекулярной биологии
.pdfПрименение чувствительных счетчиков фотонов Гей- гера-Мюллера для изучения излучения ультрафиолетовой хемилюминесценции, возникающей при различных химических реакциях (Audubert, 1938, 1939), дало большой достоверный материал, подтверждающий основные факты, полученные митогенетическими методами. Рассматривая механизм возникновения излучения, автор связывает его в основном с рекомбинациями свободных радикалов и атомов. Оценка интенсивности излучения (несколько тысяч фотонов с 1 см2 источника в 1 сек) аналогична данным, полученным другими исследователями, и соответствует квантовому выходу в 10-12—10-13 фотонов на 1 молекулу или обратно — возникновению одного фотона на 1012—1013 прореагировавших молекул.
Свободнорадикальный механизм хемилюминесценции рассматривается в настоящее время как основной. Свободные радикалы возникают при излучении окислительных процессов углеводородов в жидкой фазе и окислении липидов в модельных опытах и в живых системах.
Основной смысл |
выводов первой |
группы |
работ |
(P. Ф. Васильев, А. |
А. Вичутинский, |
1962; P. |
Ф. Ва- |
сильев, Аллабуттаев, А. А. Вичутинский, Русина, 1965), основанных на большом экспериментальном материале, следующий: окисление углеводородов в жидкой фазе представляет собой цепную радикальную реакцию; цепной процесс ведется углеводородными и перекисными радикалами; свечение реакции идет за счет энергии освобождающейся при рекомбинациях радикалов. Общий выход хемилюминесценции при этих процессах равен приблизительно 10-8 фотонов на одну образующуюся молекулу, т. е. интенсивность видимого излучения превышает на несколько порядков интенсивность митогенетического излучения, квантовый выход которого оценивается, как говорилось выше, в 10-12—10-13 фотонов на одну молекулу.
Вторая группа исследователей (Б. H. Тарусов и др., 1961, Б. H. Тарусов, А. И. Журавлев, 1965), изучающих видимую хемилюминесценцию, связанную с окислением липопротеидов, рассматривает акт высвечивания как результат взаимодействия естественных антиокислителей с перекисными радикалами, присутствующими в биолипидах организмов. Биологический смысл таких
20
взаимодействий авторы видят в постоянном поддерживании этим путем низкого уровня окисления биолипидов, особенно липопротеиновых комплексов.
Изучение коллоидных процессов и процессов кристаллизации (А. И. Рабинерсон, M. В. Филиппов, 1939, А. И. Рабинерсон, M. А. Владимирская, 1939; А. И. Рабинерсон, 1940) показало, что как при процессах коагуляции, так и при образовании трудно растворимых осадков, т. е. образовании ионных и молекулярных решеток, возникает ультрафиолетовая хемилюминесценция митогенетической интенсивности. Если для коагуляционных процессов и для процесса формирования молекулярной решетки вопрос о возникновении больших квантов энергии не является еще и в настоящее время достаточно ясным, то сам факт излучения, зарегистрированного параллельно биологическим методом и счетчиками Гейгера-Мюллера, представляет большой интерес.
Изучаемое в настоящее время (Сафонов, Шляпинтох, Энтелис, 1964) с помощью фотоэлектронных умножителей явление кристаллолюминесценции, т. е. люминесценции, сопровождающей возникновение кристаллической решетки, захватывающей видимую и близкую ультрафиолетовую область, подтверждает значение и интерес прежних данных.
Ограничиваясь этим сравнительно кратким изложением физико-химических основ митогенетического излучения, мы хотим со всей определенностью подчеркнуть несомненность тесной связи между различными подходами к изучению этого широко распространенного явления и принципиальное значение митогенетических исследований, в которых, в сущности говоря, впервые был широко рассмотрен свободнорадикальный механизм хемилюминесценции.
СПЕКТРАЛЬНЫЙ АНАЛИЗ МИТОГЕНЕТИЧЕСКОГО ИЗЛУЧЕНИЯ И ВТОРИЧНОЕ ИЗЛУЧЕНИЕ
Приводя отдельные примеры, связанные с энергетическими схемами, мы затрагивали спектральные данные. Рассмотрим основные факты подробнее, так как спектральное разложение имеет очень большое значение для любого направления исследования, связанного
21
с излучением Мы ограничимся здесь сущностью явления, а методическую сторону разберем в отдельной главе.
Возможность спектрального разложения столь мало интенсивного излучения, каким является митогенетическая хемилюминесценция, обусловлена высокой чувствительностью биодетекторов, стимуляция клеточных делений которых может быть вызвана поглощениями отдельными клетками одного-двух фотонов излучения. Поэтому и в предшествующие годы и в настоящее время рабочим методом исследования является метод биодетекции '.
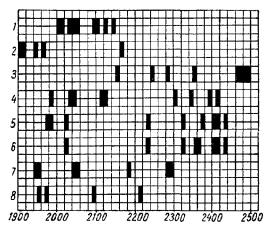
При спектральном анализе митогенетического излучения интерес может концентрироваться на изучении конечных или промежуточных продуктов реакции, т. е. стойких или мимолетных флуоресцентов, поглощающих химическую энергию и высвечивающих ее с специфическим спектром. О таких примерах мы говорили, разбирая отдельные ферментативные реакции, и таким образом был изучен ряд спектров, которые можно рассматривать как определенные ориентиры для расшифровки более сложных спектров, получаемых при известных условиях2 на биологических объектах (рис. 3).
Спектральное изучение излучения органов на живом животном при возможно близких к физиологическим условиям дает результаты, требующие другой трактовки. Вследствие диффузности спектральных максимумов выделение отдельных полос, характерных для свободных флуоресцентов, становится невозможным На первый план выступает другой критерий — оценка расположения полос и их ширины, достигающей часто значительных величин, позволяющая составить суждение о структурно-энергетическом состоянии субстрата. Этот аспект спектрального анализа подробно обсуждается в главах, посвященных анализу молекулярного субстрата живых систем и излучению нервной и мышечной систем. Здесь мы лишь подчеркнули общий характер явления.
1 Высокочувствительные фотоумножители дают возможность приблизительной оценки спектральных областей при помощи светофильтров.
2 Например, спектры излучения нервов (см. главу о излучении нервной и мышечной систем).
22
Специальной областью спектрального анализа является анализ так называемого селективного рассеяния и проведения. При этом растворы веществ (в очень малых концентрациях) или биологические объекты, подвергаясь ослабленному освещению от физического
Рис. 3. Спектры-эталоны ферментативных реакций (А. Г. Гурвич, Л Д Гурвич, 1945)
1 — расшепление креатинфосфата (является ли флуоресцентом вся молекула или продукты расщепления, осталось не вьпсненным), 2 —расщепление глюкозы (флуоресцент глюкоза), 3 —расщепление нуклеиновой кислоты и лецитина (флуоресценция группы PO4), 4 — расщепление пептидов (флуоресценция пептидной свлзи), 5 — расщепление мальтозы (флуоресцирует, по-видимому, молекула цели ком), 6 — расщепление сахарозы (флуоресцирует, по види мому, молекула целиком), 7 — расщепление мочевины (флуоресцируют радикалы R - CO и R-NH2 ), 8 — расщепление липидов (флуоресценты не ясны) По оси абсцисс —
длина волн в ангстремах
источника ультрафиолетового излучения с панхроматическим характером спектра, высвечивают ультрафиолетовое излучение митогенетической интенсивности, спектры которого состоят из различных, для каждого данного объекта, набора полос, т. е. спектры селективны.
Другими словами, в этом случае речь идет о явлении, отличающемся от хемилюминесценции в строгом смысле слова, которую именно и характеризуют спектры двух первых типов. Необходимость светового
23
возбуждения для возникновения селективных спектров, несколько нарастающая по мере распространения на большие объемы интенсивность излучения, необходимость присутствия атмосферного кислорода для возникновения излучения — все это заставляет предположить, что речь идет о сопряженных лучистых и химических процессах. Последние нужно понимать как цепные процессы разветвляющегося характера, в которых роль энергетических стартов играют фотоны. По терминологии, предложенной Я. И. Френкелем, такое явление в целом можно охарактеризовать как диффузию фотонов.
Сопряженность связи химических и физических актов принципиально важна, она лежит в основе многих наблюдаемых митогенетическими методами явлений и в живых системах обуславливает переход микроявлений в макроявления. Мы остановимся на этом вопросе подробно после описания основных результатов, полученных методом спектрального анализа селективного рассеяния. (А. Г. Гурвич, Л. Д. Гурвич, 1945, 1959).
Существенным и новым по сравнению с эмиссионными хемилюминесцентными спектрами является то, что этот метод выявляет, благодаря первичному фотовозбуждению, те элементы, которые в первом случае как бы остаются в тени. Именно так обстояло дело, как мы знаем, с обнаружением свободных радикалов при ферментативных реакциях, не обладающих без подсвечивания достаточной для митогенетического излучения энергией. Как показывает большой экспериментальный материал, «селективные» спектры делятся на два основных типа: аддитивные и интегральные. Совокупность полос в первых является как бы суммой спектров отдельных, входящих в данную молекулу и связанных с ней функциональных групп1 — гидроксильной группы, карбонильной группы, аминогруппы. Аддитивные спектры характерны для открытых цепей — спиртов, жирных кислот, аминокислот, пептидов, белков. Интегральные спектры характеризуют молекулу как целое, они типичны для циклических и гетероциклических соединений и
для |
некоторых простых |
молекул (гидроксиламина, гид- |
|
разина, |
формальдегида, |
муравьиной кислоты). |
|
1 |
Не |
являющихся, другими словами, свободными радикалами, |
|
34
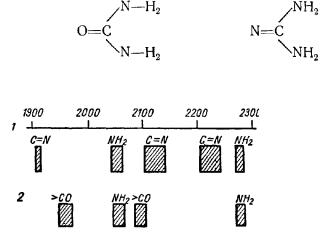
Определение спектров отдельных функциональных групп, входящих в молекулу, основывается на следующем: сравниваются спектры соединений, отличающиеся какой-нибудь одной группой. Например, сравнение
спектров мочевины |
и гуанидина |
дает предварительную ориентацию относительно того, какие полосы соответствуют аминогруппе (совпадение
Рис.. 4. Спектры селективного рассеяния растворов гуанидина и мочевины (А. Г. Гурвич, Л. Д. Гурвич, 1945).
1 —гуанилин; 2 — мочевина.
По оси абсцисс — длина волн в ангстремах.
полос) |
и какие |
карбонильной группе |
O = C |
и группе |
N = C |
(различное |
положение полос в |
обоих |
веществах |
(рис. 4).
Правильность такого вывода проверялась дальнейшим сопоставлением спектров других соединений. Так, например, полосы, предположительно характеризуемые как типичные для группы C = N, содержатся также в спектрах аргинина и креатина, но не содержатся в спектрах других аминокислот. Полосы, приписываемые аминогруппам, не содержатся в спектрах жирных кислот и т. д.
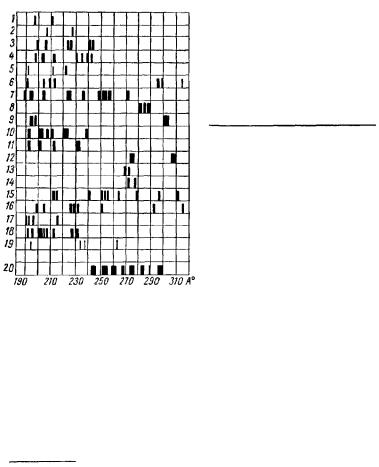
Последовательное изучение спектров целого ряда соединений позволило составить небольшой спектральный атлас, отдельные данные которого были с успехом применены для анализа молекулярных состояний субстрата живых систем (рис. 5).
25
Вернемся к очень важной связи между химическими явлениями и излучением, так как она лежит в основе возникновения митогенетических эффектов, остающихся на уровне микроявлений в неорганизованных системах, по приводящих в живых системах к различным макропроявлениям.
Возникновение макроявлений обуславливается развитием цепных процессов разветвленного характера.
Фотоны излучения являются при этом стартовыми факторами химических процессов — молекулы, возбужденные поглощенной энергией, становятся центрами заро-
Рис. 5. Спектры селективного рассеяния и хемилюминесценции различных соединений:
1/ — карбонильной группы; 2 — аминогруппы; 3 —гидроксильной группы; 4 — лактимной формы пептидной связи; 5— лактамной формы пептидной связи; 6—метальной группы; 7 —метилового спирта; 8 — гидроксиламина; Р—гидра-
зина; |
10 — формальдегида; /7 —индола, |
|
12 — пиррола; 13 — фенильной |
группы; |
|
/4 —оксифенильной группы; |
15— аде- |
|
нина, |
16 — дикетопиперазина; 17 — глю- |
|
козы; |
IS- триптофана; 19 — ацетилхо- |
|
|
лина; 20 — бензола. |
|
ждения цепей. U разветвляющемся характере цепных процессов говорит в первую очередь основной митогенетический эффект, т. е. стимуляция митогенетическим излучением клеточных делений. Действительно, деление клетки — процесс, связанный с перестройкой всего клеточного субстрата, вызывается поглощением клеткой лишь одного-двух фотонов излучения1.
На это же указывает широко распространенное явление вторичного излучения, возникающего при облучении соответствующих субстратов источниками митогенетического излучения. Вторичное излучение распро-
1 Подробное изложение фактов дается в главе, посвященной клеточному делению.
26
страняется на большие объемы и по мере распространения не только не затухает, но даже несколько нарастает в своей интенсивности.
Другими словами, облучение субстрата вызывает в нем фотохимические цепные процессы, сопровождающиеся излучением. Вторичное излучение распространяется в растворах со скоростью порядка 30 ж в 1 сек. Латентный период, отделяющий вспышку вторичного излучения от подаваемого в виде короткой вспышки первичного облучения, порядка 0,001 сек.
При облучении субстрата источником митогенетической интенсивности важно, чтобы в спектр источника входили хотя бы некоторые полосы, соответствующие спектру вторичного излучения субстрата. При облучении более интенсивным физическим источником ультрафиолетового излучения это условие не является обязательным.
Принципиально важным является тот факт, что способность к вторичному излучению характерна только для свежих или приготовленных недавно растворов. Так, например, растворы глюкозы, мочевины, нуклеи-
новых кислот, белка |
(особенно при их хранении на свету |
||
и в |
присутствии атмосферного кислорода) |
сохраняют |
|
свою |
способность |
к вторичному излучению лишь в |
|
течение нескольких |
часов. Параллельно с |
такой ла- |
|
бильностью соединений наблюдается и свойство быстрой утомляемости растворов, т. е. потери ими способности к вторичному излучению, наступающей, как правило, через 15—20 мин после начала облучения митогенетическими источниками.
Больше того, утомленный раствор обладает свойствами гасителя излучения (т. е. оптической непрозрачно-
. йости) |
по |
отношению к свежеприготовленным. Так, |
например, |
раствор глюкозы, облучавшийся длительное |
|
время, |
гасит вторичное излучение свежего раствора при |
|
прибавлении |
к нему в отношении 1 : 100. Такой же эф- |
|
факт гашения наблюдается и при прибавлении необлученных растворов, но хранившихся длительное время на свету. Эти результаты позволяют, таким образом, сделать вывод, что в обоих случаях происходят одина-
ковые химические изменения.
Вторичным излучением не исчерпываются те несомненные фотохимические эффекты, которые вызывает
27
митогенетическое облучение объектов, но во всех случаях поглощение фотона должно быть стартом цепной реакции с выходом во всяком случае больше единицы.
Другими словами, первичный элементарный акт приводит к большему числу вторичных актов, а при облучении живых систем к значительно большему числу. Мы подчеркиваем эту формулировку, так как именно в митогенетических исследованиях анализируется вопрос о роли редких событий в развитии биологических явлений, вовлекающих несоизмеримо большее число молекул, по сравнению с активизированными вначале. Специфические условия молекулярного субстрата живых систем, благоприятствующие развитию процессов, будут поэтому рассматриваться весьма подробно.
Цепные реакции, доступные нашему наблюдению, разделяются во-первых, на реакции, сопровождающиеся излучением (вторичное излучение) и протекающие без излучения; во-вторых, они различаются по характеру конечных продуктов на реакции с образованием продуктов фотодиссоциации и реакции синтетического характера.
Все полученные до сих пор результаты говорят о том, что реакции, сопровождающиеся излучением, относятся к процессам диссоциаций, а реакции, не сопровождающиеся излучением, являются реакциями синтеза. В сложных субстратах живых систем, несомненно, осуществляется сопряженность этих явлений.
РЕАКЦИЯ СИНТЕЗА
Разберем вопрос о поликонденсации пептидов, обнаруживаемой в модельных опытах. Конденсация связана с установлением связи между мономерами путем выделения молекулы воды:
НО—CH2-ОН+НО—CH2-OH —> ОН—CH2 -О—CH2 -ОН+Н2 О
При обычных условиях воздействия, например нагреве, конденсация не представляет собой цепного процесса, так как совокупность актов, приводящих к формированию пептидной связи, приблизительно термонейтральна, т. е. при этом не освобождается энергия для построения дальнейшей пептидной связи. При воздействии фотонами митогенетического излучения процесс
28
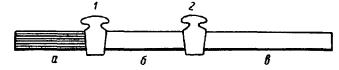
приобретает цепной характер. Это является понятным, так как поглощенная аминокислотой достаточно высокая энергия фотона, дающая начало процессам перестройки, освобождается при завершении этих актов и может быть использована для построения следующей пептидной связи. Другими словами, при облучении и небольшой непрерывной энергии активации процесс конденсации может распространиться, несмотря на известный градиент падения, на измеримые объемы
Рис. 6. Трубка с кранами (А. Г. Гурвич, Л. Д. Гурвич, 1945).
Объяснение в тексте
(А. Г. Гурвич, Л. Д. Гурвич, 1945, 1959). Это подтверди дается экспериментально. Трубка длиной в 20 см, разделенная кранами на три отдела, заполнялась раствором пептона. Во время короткого (1—2 мин) облучения пептона в отделе «а» отделы «а» и «б» оставались сообщенными, отдел «в» был отделен. После облучения
отдел «б» отделялся от «а», и |
через |
10 мин на корот- |
кое время открывался отдел |
«в». |
Примерно через |
30 мин из отдела «в» бралась проба пептона, которая при соединении с пепсином (желудочный сок) давала вспышку митогенетического излучения. Присоединение пепсина к необлученному пептону не давало излучения (рис. 6). Таким образом, цепной характер поликонденсации является несомненным.
Наряду с пептоном, облучению подвергались с положительным результатом и смеси аминокислот — дикарбоновые кислоты и гликокол, иногда включалась и циклическая аминокислота (тирозин).
Образующиеся в очень малых концентрациях конденсаты высокомолекулярны, на что указывает их неспособность к диализу через коллодийную пленку. При прибавлении пепсина к отделенному диализом синтетическому продукту возникает митогенетическое излучение со спектром, характерным для пептидной связи.
29