

Проблема митогенетического излучения как аспект молекулярной биологии
.pdfобразов, А. Г. Гурвич писал: «Наши зрительные образы, наряду с абсолютно обратимыми, однозначно связанными с содержанием зрительных полей впечатлениями, содержат и необратимые слагаемые, часть которых во всяком случае не только не является однозначной функцией от внешних полей зрения, но и не совпадает с ними по содержанию. Обучение, проторение, память относятся к категории необратимых явлений. Существуют геометрические конфигурации, не адекватные для органа зрения, в таких случаях они заменяются другими, искаженными (геометрически — оптические обманы зрения)».
«Наряду с этим, из простых рассуждений вытекает, что гистерезис, свойственный нашим зрительным восприятиям, не связан с теми инстанциями зрительного аппарата, которые обуславливают возникновение элементарных ощущений. Например, очевидно, что простейший случай гистерезиса — узнавание зрительного образа при его вторичном появлении происходит и тогда, когда в обоих случаях картина захватывает различные элементы зрительного эпителия (например, последовательное восприятие двух подобных треугольников различной величины).
Мы должны таким образом принять, что гистерезис может быть обусловлен в этом случае, как и в бесчисленном множестве других, только той инстанцией, которая является в какой-то степени автономной относительно аппаратов, связанных с элементарными возбуждениями, т. е. соответствующих нейронов.
Мы можем охарактеризовать эту инстанцию как «континуум», т. е. как тот основной субстрат, в который как бы «погружены» морфологически определимые элементы: клетки, волокна. Другими словами, мы не отождествляем его с нейропилем» (А. Г. Гурвич, 1929).
Понятие неравновесной молекулярной упорядоченности не только гармонирует с понятием континуума, но делает доступным его экспериментальное изучение. Значительную роль сыграло при этом изучение митогенетического излучения коры при зрительном раздражении. Можно выделить две группы результатов: во-первых, изложенные уже коротко в начале этой главы данные, показывающие, что локализованное и неглубокое поранение поверхности мозговых полушарий лягушки при-
150
водит к далеко идущим последствиям при одновременном освещении глаз. Возникающий при этих условиях митогенетический шок, т. е. временная потеря способности к излучению, захватывает не только полушарие целиком, но распространяется на всю зрительную систему, насколько об этом можно судить по хиазме зрительного нерва. Наиболее правдоподобное объяснение заключается в том, что зрительное возбуждение объединяет всю зрительную систему в одно общее функциональное целое.
Подтверждение и некоторый анализ этого вывода стали возможными на основании второй группы фактов. Были сопоставлены результаты, полученные при изучении излучения коры нормальных кроликов и кроликов, находящихся в состоянии экспериментально вызванной каталепсии (внутривенное введение небольших доз гистамина) (С. H. Брайнес, 1934, 1937), т. е. значительном отклонении нервных центров от нормы. У обоих групп животных излучение изучалось при различных функциональных состояниях и при охлаждении коры.
Полученные данные позволяют выделить два типа состояний континуума:
1) при возможном выключении внешних раздражителей, но при непрерывной, по-видимому, смене разнообразных состояний внутреннего происхождения континуум нужно рассматривать как совокупность различных неравновесных молекулярных констелляций. Тот же характер континуума сохраняется и при применении тактильных или болевых раздражений (слабой электризации кожи). Ближе всего такое состояние можно обозначить термином «потенциальный» континуум;
2) при световом и звуковом возбуждениях, т. е. адекватных раздражениях сложных и совершенных рецепторов состояние молекулярного континуума коры изменяется. Констелляции объединяются общими энергетическими уровнями на больших протяжениях. Схематически говоря, происходит нечто вроде возникновения очень обширного трехмерного неравновесного молекулярного ансамбля. Континуум такого характера назовем «актуальным».
Говоря об объединении континуума в одно целое, мы широко используем представление о непрерывности молекулярного субстрата, не ограниченной разделами, обусловленными поверхностями клетки или волокна. С точки зрения концепции поля это является
151
вполне естественным, мы помним, что сфера действия клеточного поля выходит за границы клетки. Именно путем геометрического сложения клеточных векторов создается актуальное поле совокупности клеток.
Само собой разумеется, что представления о континуумах обоих типов сохраняют чисто динамический характер, т. е. речь идет о поддержании, непрерывном воспроизведении пространственных неравновесных молекулярных структур или молекулярной архитектуры.
Экспериментальные результаты, обосновывающие эти представления, заключаются в следующем. При спокойном состоянии, т. е. возможном выключении внешних раздражений, характерными являются довольно большая длительность деградационного излучения, достигающая при продолжающемся охлаждении до 6—8° 10—15 мин, и изменение спектра деградационного излучения во времени.
Тот же характер деградационного излучения обнаруживается и при охлаждении коры кроликов, находящихся в состоянии каталепсии. Более детальное сравнение этих двух состояний, при котором могли бы выявиться некоторые различия, пока еще не сделано.
Таким образом, несмотря на несомненное различие функциональных состояний коры у нормальных и каталептических кроликов, для них свойственно и очень существенное общее: констелляции молекулярного континуума сравнительно мало ограничены в своих параметрах, степени свобод в распределении энергетических состояний, а следовательно, и конфигураций констелляций, сравнительно велики, т. е. констелляции не объединены в одно энергетическое целое. Именно как следствие постепенности нарушений различных констелляций можно объяснить и длительность деградационного излучения и изменения спектров во времени.
Эти результаты подтверждают прежние данные о том, что физиологическое излучение коры (при устранении раздражений) нужно рассматривать как проявление непрерывных нарушений отдельных, качественно различных, неравновесных молекулярных ансамблей, возникновение которых является выражением непрерывной смены состояний внутреннего происхождения.
Дальнейшие опыты с применением болевых и тактильных раздражений расширили это представление.
152

Выяснилось, что раздражения, усиливая у нормальных и каталептических кроликов и спонтанное излучение коры, и деградационное излучение (при одновременном с раздражением охлаждении коры), не изменяют вместе с тем длительности излучения. Она достигала при одновременном раздражении и охлаждении тоже 10— 12 мин. Таким образом, при большем числе нарушений констелляций в единицу времени (что вызывает усиление излучения), т. е. при большей частоте формирования констелляций, общим с состоянием покоя остается энергетическое и конфигурационное разнообразие констелляций. Вместе с тем, нужно, очевидно, допустить, что какая-то специфика конфигураций отражает состояние возбуждения, вызванного раздражением кожных рецепторов.
Другими словами, континуум при этих раздражениях представляет также совокупность отдельных неравновесных молекулярных систем, т. е. соответствует термину «потенциальный».
Другая и, по-видимому, принципиально отличная картина выясняется при сопоставлении ряда данных, в которые входят и результаты, получаемые при зрительном и звуковом раздражениях. Кора нормальных кроликов реагирует на световое и звуковое раздражение усилением излучения. Освещение глаз у каталептических кроликов не приводит к усилению излучения.
Вторичное излучение1 распространяется при световом возбуждении на большие протяжения в коре нормальных кроликов и быстро затухает при этих же условиях в коре каталептических кроликов. Этот факт ясно указывает, что в то время как в норме световое возбуждение способствует распространению цепных процессов, при каталепсии возникают условия, препятствующие их распространению.
Наряду с этим, различие результатов наблюдается у нормальных и каталептических кроликов при совмещении светового возбуждения коры и охлаждения ее поверхности. Излучение коры нормального кролика при этих условиях резко снижается, излучение коры каталептических кроликов держится приблизительно на том
1 Локальное митогенетическое облучение небольшого участка коры приводит к распространению излучения на большую поверхность. Это явление можно рассматривать только как распространение цепных процессов, связанных с высвечиванием энергии.
153
же уровне, как при охлаждении мозговой поверхности без светового возбуждения.
Сопоставление всех этих данных приводит нас к тому представлению, которое раньше было высказано в виде предположения. Адекватные раздражения у нормальных животных таких сложных рецепторов, как зрительный и звуковой, приводят молекулярный субстрат коры (специфических зон, а может быть и более обширных областей) к сдвигу динамического равновесия в сторону образования протяженной системы общих энергетических уровней. С этой точки зрения становится вполне понятным хорошее проведение вторичного излучения при световом возбуждении коры.
Потерю способности к излучению при одновременном световом возбуждении и охлаждении коры правдоподобнее всего объяснить тем, что деградация протяженных неравновесных систем приводит не к их распаду, т. е. возникновению флуоресцентов, а напротив, сдвигает равновесие в сторону образования еще большего объединения и несколько большей равновесности молекулярных систем, способность к флуоресценции которых понижена.
Другими словами, выражение возбуждения коры при зрительном (и, по-видимому, при звуковом) раздражении мы видим именно в возникновении «актуального» молекулярного континуума. Причем нам представляется очень вероятным, что акт энергетического объединения констелляций, приводящий к возникновению определенной молекулярной архитектуры, является специфической, т. е. ответственной за возникновение данного зрительного ощущения, фазой.
С этой точки зрения проявление состояния каталепсии можно видеть в «блокировании» молекулярного континуума на отдельные области, препятствующем возникновению общей системы энергетических уровней и, вследствие этого предположить, что зрительные восприятия животных при экспериментальной каталепсии сравнительно элементарны.
Говоря о возникновении актуального континуума мозговой коры при световом возбуждении, мы связываем представление о нем с разнообразием стерических параметров, которые должны находиться в какой-то функциональной зависимости от характера световых раздражений.
154
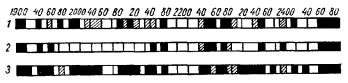
Конкретные результаты дает в этом отношении спектральный анализ (А. А. Гурвич, 1934). На интактных лягушках и на отрезанной голове лягушки (в течение первого часа после декапитации) были изучены спектры излучения полушарий, зрительных долей и зрительного нерва (в области перекреста) при довольно ярком панхроматическом освещении глаз.
Спектры этих трех областей значительно отличаются друг от друга (рис. 25). Наряду с этим сравнивались спектры полушарий при освещении глаз различными цветными полями зрения. Несмотря на сравнение только отдельных спектральных областей, специфиче-
Рис. 25. Спектры излучения возбужденной зрительной системы лягушки (освещение глаз) (А. Г. Гурвич, Л. Д. Гурвич, 1945).
7 —полушария; 2—зрительные доли; 3—зрительный нерв (область перекреста).
ская зависимость спектров от данного монохроматического освещения выражена ясно.
Значительно большее разнообразие данных было получено при помощи метода, применявшегося уже к периферическим нервам и основанного на изучении распределения интенсивности излучения мозговой поверхности лягушки и кролика. Отчетливые результаты получались при выделении с помощью специального приспособления какой-нибудь слагаемой из общего, несомненно богатого и должно быть широкополосного спектра, например флуоресценции пептидов или глюкозы.
При проецировании на роговицу глаза полей зрения, состоящих из различных комбинаций освещенных и затемненных частей, были зарегистрированы как различия, так и совпадения кривых распределения интенсивности (А. А. Гурвич, 1938). Освещение верхней половины глаза и затемнение нижней и обратно, затемнение верхней и освещение нижней давали кривые излучения одинакового характера. Замена поля зрения, содержащего темный зигзаг на светлом фоне, полем со светлым
155
зигзагом на темном фоне приводила к изменению кривых излучения. Замена же темного квадрата на светлом фоне темным кругом на светлом фоне не отражалась на кривых. В ряде опытов было замечено, что скользящий перед глазом лягушки темный рисунок на светлом фоне не изменял кривой излучения по сравнению с исходной кривой, соответствующей светлому полю зрения, в то время как такой же неподвижный рисунок изменял характер кривой (рис. 26).
Эти немногочисленные и поэтому ориентировочные данные интересны тем, что, показывая на разнообразие параметров излучения как симптома возбужденных состояний, они вместе с тем указывают и на определенную автономность мозговой коры, в смысле «неподатливости» относительно некоторых раздражений.
Мы возвращаемся, таким образом, снова к представлению о континууме и попытаемся в связи с этим изложить возможно яснее те представления А. Г. Гурвича, в которых общее понятие молекулярного континуума синтезируется с представлениями, развивающимися на клеточном уровне, т. е. связывается с понятием актуального поля мозговой коры.
«В нашей попытке анализа мозговой коры мы будем исходить не из ее элементаризирующего расчленения, но, наоборот, из выделения основных данных, характеризующих ее как целое.
Одним из основных фактов является то, что число клеток в участке коры, соответствующее, например, данному рецептору, значительно больше числа нервных волокон, связывающих их с элементами рецептора. Чрезвычайное богатство и густота разветвлений делают, с одной стороны, неизбежным заключение, что все клетки оплетаются афферентными волокнами, но исключают, с другой стороны, всякую вероятность изолированной, т. е. нарочитой, связи между одним определенным волокном и определенной группой клеток. Не имеет поэтому смысла говорить о функции клетки коры, взятой в отдельности, т. е. в противоположность клеткам спинного мозга и может быть низших центров, клетки коры нельзя назвать функциональными единицами и принять простую аддитивность их функционирования.
Замечательна также архитектурность мозговой коры — закономерное пространственное распределение и
156
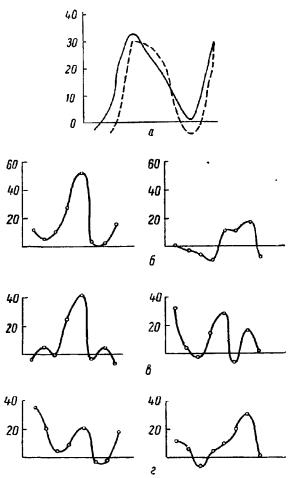
Рис. 26. Распределение интенсивности излучения мозговой коры кролика при различных зрительных восприятиях (А. Г. Гурвич, Л. Д. Гурвич, 1959).
а —освещение верхней половины |
глаза (сплошная линия) |
и нижней половины (пунктирная); |
б — светлое поле зрения; |
в — подвижная линия на светлом поле (слева), неподвижная линия на светлом поле (справа); г — неподвижная линия на светлом поле (слева), подвижная линия (справа).
По оси ординат — эффект в процентах.
157

ориентировка входящих в ее состав клеток. Такая архитектурность является прерогативой лишь коры полушарий и мозжечка. Этот факт делает логически неизбежным вывод о существовании между клетками, взаимоотношений, охарактеризованных пространственными (геометрическими) параметрами. При этом наблюдается типичное своеобразие конфигураций различных категорий клеток коры и максимальное развитие их поверхности в виде отростков, также охарактеризованных определенными для каждой категории параметрами. Богатство разветвлений относится в равной мере и к приходящим в мозговую кору из низших центров нервным окончаниям. Необходимо при этом отдать себе отчет в том, что весь характер разветлений нервных клеток лишь типичен для каждой из них, но флуктуирует в очень широких пределах, подобно тому как лишь типична ветвистость деревьев». Другими словами, характер разветвлений нормирован1.
«Наличие хроматина, помимо ядра, и в цитоплазме самого тела клетки и в ее разветвлениях (дендритах) позволяет дать клеткам, с точки зрения поля, следующую характеристику: а) каждая нервная клетка является источником поля и притом, по-видимому, интенсивным (энергичный метаболизм хроматина); б) геометрические параметры собственного поля каждой клетки являются функцией от ее конфигурации, включая в это понятие и отростки (поскольку они содержат хроматин)
Конструкция актуального поля (в своей самой общей форме) исходит, как известно, из того, что сфера действия клеточного поля выходит за пределы клетки и, что следовательно, хотя вектор поля и убывает в какой-то обратной зависимости от расстояния, в любой точке кле-
1 Принцип нормировки, как уже говорилось, органически связан с статистическим подходом.
В связи с этим интересно привести взгляды Sholl (1956). Считая невозможным в настоящее время создание общей удовлетворяющей модели организации коры, он говорит- «Тем не менее, мы все употребляем модели и в связи с этим новый язык, язык статистики, основывающийся на вероятностных концепциях, должен быть предложен». «Предполагается, что необходимость статистического подхода к изучению коры обуславливается не только шириной и сложностью экспериментальных наблюдений, но что она связана с самой природой организации коры, системы, которая может быть описана только в понятиях теории вероятности».
158

точного комплекса существует поле, вектор которого определяется полями всех входящих в состав комплекса клеток. Он обозначается как вектор макрополя. Архитектура макрополя определяется рядом параметров — числом клеток, их пространственным распределением и характером их собственных полей. Строгая архитектоника мозговой коры — однородность строения и конфигураций преобладающих в данной области коры элементов (пирамидные клетки), протяженность их распространения и, особенно, строго параллельная ориентировка их осей — приобретает поэтому глубокий смысл. Однако вследствие особенностей отдельных цитоархитектонических участков коры рационально относить понятие макрополя к этим отдельным участкам. Но, вместе с тем, и в этом и заключается ценность полевой концепции, вполне реально представление об общем поле мозговой коры: моментальное состояние его любого участка отражается и на состоянии отдаленных областей»1.
Широкое распространение цитохроматина в дендритах и богатстве их разветвлений, т. е. большая эффективность актуального поля, позволяет говорить о значительной степени векторизации молекулярных процессов в структурных элементах коры и вне их. Другими словами, молекулярный субстрат коры выраженно неравновесен и мы помним, по данным митогенетического анализа, как легко совершается переход потенциального континуума в состояние актуального, в котором понятие неравновесности относится к большим областям коры.
Приведем некоторые выдержки из физиологической литературы, указывающие на аналогию в подходах к анализу функционирования нервных центров и мозговой коры как целого.
«... Надо смело признать, — говорит П. К. Анохин (1935), — что нервная система осуществляет свою интегративную деятельность не только по структурным, но
и по специфически-функциональным принципам, которые
уформирующейся нервной системы имеют превалирующее значение. Этим и обуславливается то динамическое единство специфичности и целостности, с которыми мы встречаемся при изучении нервной деятельности у взрослого животного».
1Из неопубликованных трудов А. Г. Гурвича.
159