

Проблема митогенетического излучения как аспект молекулярной биологии
.pdfТ А Б Л И Ц А 18
Спектральный анализ излучения икроножной мышцы
|
Спокойное |
излучение |
Деградационное |
излучение |
|||||
Длины волн в Å |
МЫШЦЫ В % |
|
|
мышцы в % |
|||||
|
|
|
|
|
|
|
|
|
|
|
лягушки при |
кролика при |
лягушки при |
|
кролика при |
||||
|
15-16° С |
35-38° С |
2-3° |
|
10-12° |
||||
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
1900—2000 |
— 3 |
— 7 |
2 |
|
— 3 |
||||
2000—2100 |
1 |
— 1 |
3 |
|
7 |
|
|||
2100—2200 |
5 |
1 |
12 |
|
0 |
||||
2200-2300 |
50 |
53 |
42 |
|
55 |
||||
2300—2400 |
— 7 |
2 |
5 |
|
— 3 |
||||
2400—2500 |
2 |
— 3 |
|
|
2 |
|
|
|
|
2500—2600 |
1 |
- 6 |
11 |
|
4 |
||||
2600—2700 |
5 |
— 3 |
1 |
|
|
|
2 |
||
2700—2800 |
- 2 |
2 |
4 |
|
6 |
||||
2800—2900 |
— 1 |
5 |
8 |
|
3 |
||||
2900—3000 |
4 |
— 6 |
|
— 2 |
|
9 |
|||
3000—3100 |
5 |
1 |
13 |
|
2 |
||||
3100—3200 |
2 |
8 |
1 |
|
11 |
||||
|
|
|
|
|
|
|
|
|
|
расположен дальше от коллиматора спектрографа, чем лягушка. Этим объясняется приблизительная идентичность экспозиций, несмотря на более интенсивное излучение мышц теплокровных по сравнению с лягушками.
Грубое разложение спектров показывает не только несомненное сходство мышц лягушки и кролика, но и сходство спокойного и деградационного излучения Это дает уже значительно более веское обоснование представлению о физиологическом состоянии спокойной мышцы как о естественно протекающих сменах нарушений и восстановлений неравновесной молекулярной упорядоченности.
Более детальное изучение эффективной области спектра (разложение 2200—2300 Å), проделанное пока для спокойного состояния икроножной мышцы, показало реальную ширину спектральной полосы, равную в среднем 60—70 Å. При повторении опытов наблюдалась, однако, несомненная флуктуация границ. Этот последний факт вполне естествен, указывая на лабильность излучающего субстрата. Вместе с тем, он позволяет дать только приблизительное определение положения полосы, занимающей область в указанных границах или выходящей
120

несколько за их пределы. Различия между спектрами лягушки и кролика не выходят за пределы этих флуктуации.
Обнаруженная ширина полосы представляет, с нашей точки зрения, большой интерес Мы увидим в дальнейшем, что степень ширины спектральных полос можно, по-видимому, рассматривать как критерий молекулярной упорядоченности саркоплазмы.
Вместе с тем, бросается в глаза большая ограниченность эффективной области спектра икроножной мышцы, выраженной только одной полосой Естествен был вопрос, не является ли такая ограниченность характерной для спокойного излучения и других мышц. Изучение спектров излучения полуперепончатой мышцы лягушки и двуглавой мышцы кролика показывает, однако, что для этих мышц характерно большее число полос, большинство из которых обладает тоже довольно значительной шириной. Распределение их для каждой мышцы различно и специфично.
Такая «спектральная индивидуальность» спокойных мышц приводит, как нам кажется, к следующему важному выводу: одинаковое, с функциональной точки зрения, состояние покоя различных мышц осуществляется при различных вариантах их молекулярных субстратов, т. е. с молекулярной точки зрения состояния покоя различны. Как связать эти два положения?
Перед нами встает следующая альтернатива: молекулярные представления ничего не дают для анализа субстрата возбудимости. Или, напротив, молекулярные представления полезны, заставляя нас принципиально расширить характеристику физиологического состояния покоя систем !.
Необходимо ввести понятие функционального соответствия или функциональной адекватности систем друг относительно друга, т. е. рассматривать их всегда с точки зрения непрерывно осуществляющегося взаимодействия между ними.
С молекулярной точки зрения это означает, например, наличие общих параметров в организации субстратов систем, дающих возможность распространения более
1 Понимая системы в самом общем смысле — нервные центры, рецепторы, двигательные системы.
121
или менее подобных стерических цепных процессов на достаточно большие протяжения.
Анализ непрерывного и глубокого, т. е. относящегося к молекулярному уровню, взаимодействия отдельных систем внутри обширной функциональной системы как целого будет задачей всего дальнейшего изложения.
Излучение икроножной мышцы в различные возрастные периоды животного. Ясное подтверждение изложенного выше дает группа данных, полученных при изучении излучения икроножной мышцы маленьких кроликов (А. А. Гурвич, В. Ф. Еремеев, M. А. Липкинд, 1961).
Опыты ставились на животных, начиная с односуточного и до двухнедельного возраста. Нужно отметить, что к мышцам кроликов в первые дни после рождения неприменимо, в сущности говоря, понятие покоя в том смысле, как к взрослым животным. Это видно и по почти непрерывной подвижности маленьких кроликов и известно из электрофизиологических данных. Для экспозиций, во время которых регистрировалось излучение, выбирались поэтому те короткие промежутки времени, в
Т А Б Л И Ц А 19
Интенсивность спокойного и деградационного излучений икроножной мышцы кроликов в различные возрастные периоды
|
|
Эффект |
Экспозиция |
Деградапиои- |
Возраст в |
Экспозиция |
спокойного |
ное |
|
сутках |
в секундах |
излучения в |
в секундах |
излучение в |
1—2 |
3 |
40 |
5 |
5 |
||
|
|
|
8 |
36 |
||
|
3 |
|
Ю |
|
|
2 |
4 - 5 |
—3 |
СО |
|
|
||
|
||||||
|
5 |
28 |
|
27 |
||
|
3 |
|
ю |
|
|
|
9 |
10 |
со |
—10 |
|||
|
5 |
36 |
|
20 |
||
|
|
|
|
|
|
|
16 |
Сл |
2 |
3 |
48 |
||
|
OO |
24 |
|
|
|
|
|
|
|
|
|
|
|
Взрослые кро- |
5 |
—3 |
3 |
39 |
||
лики |
8 |
45 |
|
|
|
|
П р и м е ч а н и е . |
Приведены средние величины из двух опытов. |
|
|
|
||
122
течение которых иммобилизированные на станке животные были во всяком случае внешне спокойны.
Было показано, что спокойное излучение, выраженное наиболее ярко в первые дни постнатального развития, убывает по мере роста животного и после двух недель становится по интенсивности приблизительно стабильным.
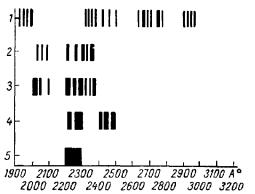
Рис. 19. Спектры излучения икроножной мышцы кролика в различные возрастные периоды (А. А. Гурвич, 1966).
1 — 1-е; 2 — 2-е; 2 — 6-е; 4 — 9-е; 5 —16-е сутки.
Изменения деградационного излучения имеют обратный характер, т. е. с ростом животных наблюдается нарастание интенсивности, приобретающей приблизительно тоже к двухнедельному возрасту постоянный характер (табл. 19).
Паралльно с этим происходит эволюция спектров спокойного излучения мышцы. Большое количество узких полос, характерное для первых дней развития, постепенно убывает; возникающие новые полосы, группирующиеся главным образом в более коротковолновой части спектра, характеризуются большей шириной, которая по мере увеличения возраста становится все более выраженной. К двухнедельному возрасту спектр приобретает свой окончательный, типичный для взрослого животного, характер — он содержит одну широкую полосу в области 2200—2300 Å (рис. 19).
123

К этому же сроку заканчивается формирование мионейральных связей, вырабатывается поза стояния и наступает прозревание животного, т. е. возникновение полноценного функционального состояния высших центров.
Возможность сопоставления митогенетических и функциональных данных и выявления параллелизма между ними является, конечно, важной, показывая еще раз физиологичность наблюдаемых молекулярных явлений и необходимость изучения более глубоких связей1. Но, помимо этого, из полученных результатов можно было сделать и более частные выводы: 1) широкополосность спектров является показателем возникновения неравновесной молекулярной упорядоченности; 2) результаты еще раз указывают (и на этот раз без экспериментального вмешательства) на значение центральных воздействий для непрерывного поддерживания неравновесной молекулярной упорядоченности субстрата мышц взрослых животных.
Излучение мышцы при ослаблении воздействия со сто-
роны центров. Дальнейшее изучение непрерывной «настройки» молекулярного субстрата саркоплазмы, идущей со стороны нервных центров, проводилось при нарушении или ослаблении воздействия центров:
1)спектр излучения мышцы изучался через некоторое время после перерезки мозгового ствола у основания продолговатого мозга, т. е. нарушения целости нервной системы в отдаленном от мышцы участке;
2)во время общего эфирного наркоза, тормозящего
впервую очередь активность высших центров;
3)во время охлаждения значительного участка спинного мозга через костный покров, т. е. без нарушения целости позвоночника, но при удалении спинных мышц.
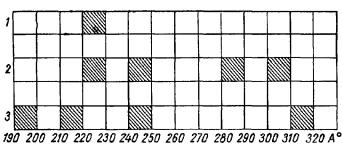
Результаты, полученные при этих трех модификациях, аналогичны. Вместо одного широкого максимума излучения (2200—2300 Å) возникает ряд полос в различных участках спектра (рис. 20). Судя по еще незаконченным
1Несомненный интерес представляет, с этой точки зрения, сопоставление наших данных с детальными исследованиями нервномышечного аппарата в ранние возрастные периоды животных, проводимыми И А. Аршавским и его школой (Аршавский, 1958. Розанова, 1938, 1941).
124
данным детального расчленения спектров, ширина большинства полос значительно меньше.
Наряду с этим спектр излучения икроножной мышцы изучался также и после длительного общего охлаждения лягушек (двухсуточное сидение на льду). Этим путем можно было рассчитывать получить сведения о тех вероятных изменениях неравновесного молекулярного субстрата мышцы, которые связаны с понижением метаболизма всего организма. Причем нельзя было исключить
Рис. 20. Спектры излучения икроножной мышцы лягушки (А. А. Гурвич, 1962).
1— в норме; 2 — при обшем охлаждении; 3—при общем наркозе.
возможность того, что такое длительное постепенное понижение метаболизма может быть связано не только с количественным изменением энергетического уровня, но и с изменением характера протекающих процессов — меньшей вероятностью глубоких диссоциаций молекул, меньшей возможностью синтезов и т. д.
Изменения спектра излучения проявлялись при этом условии в возникновении ряда новых полос, причем, судя по полученным данным, они тоже значительно сужены.
Таким образом, и нарушение, и ослабление центральных воздействий, и понижение общего энергетического уровня организма приводят, по-видимому, к значительному ослаблению степени упорядоченности субстрата мышцы. Сужение спектральных полос и их разбросанность по всему спектральному диапазону, несомненно, связано с этим.
125
ЗАВИСИМОСТЬ ИЗЛУЧЕНИЯ НЕРВОВ ОТ ФУНКЦИОНАЛЬНОГО СОСТОЯНИЯ МЫШЦ
Мы подробно рассмотрели явления, показывающие значение непрерывного регулирующего воздействия нервных центров для поддержания молекулярной упорядоченности саркоплазмы мышц, но митогенетическое излучение показывает, что понятие одностороннего воздействия является недостаточно полным.
Наши представления о нервной и мышечной системах должны быть связаны с принципом взаимодействия в непрерывном и полном смысле этого слова, означающем, что «организация» молекулярного субстрата мышечных и нервных элементов является в каждый данный момент в значительной степени результирующей их взаимодействия.
Экспериментальный материал, полученный на периферических нервах, показывает, что и смешанные нервные стволы (седалищный нерв) и чисто чувствительные нервы излучают только в том случае, если они входят в состав более обширной и сложной системы, т. е. если сохраняется их связь с центрами или периферией, а для некоторых состояний и с центрами и с периферией. Это последнее является обязательным для состояния покоя (тонуса) и для состояний возбуждения, вызванных адекватными раздражениями рецепторов.
Излучение, возникающее при искусственных приложенных непосредственно к нерву раздражениях, не требует сохранения связи и с центром, и с периферией. Причем, в этих случаях наблюдается различие между смешанными и чисто чувствительными нервами Для излучения седалищного нерва необходимо и достаточно сохранение связи с мышцами. Напротив, для излучения чисто чувствительных нервов, например спинных кожных нервов лягушки, необходимой является связь с центрами, периферический конец нерва может быть нарушен
Нарушение связи нервов с центром и с периферией на седалищном нерве и на кожных нервах (лягушка) достигалось различными путями. Опыты, производившиеся через некоторое время после нанесения травмы, ставились как на спинальных лягушках, так и на нервномышечных препаратах (с сохранением спинного мозга
126
и всех мышц голени и нижней части лапки) Эффект излучения седалищного нерва лягушки составлял:
|
% |
В норме |
26,24,20,29; |
После перерезки дистального |
|
конца |
6, 8, |
»» . проксимально-
го конца |
11, 6. |
Не анализируя пока результатов глубже, мы можем все же сказать, что излучение нерва, отражающее полноценное физиологическое состояние покоя, связано с непрерывным активным взаимодействием центров и периферии При рефлекторном раздражении (фарадизация контралатерального нерва) излучение наблюдается тоже при сохранении связи только с мышцей (табл 20). Связь с мышцей обуславливает также способность нерва к проведению вторичного излучения.
Т А Б Л И Ц А 2 0
Зависимость излучения нерва от различных воздействий |
||||||
|
|
|
|
|
|
|
|
|
|
|
|
Эффект излучения в % |
|
|
|
|
|
|
при нарушении целости |
|
|
Способ раздражения |
|
|
|
||
|
|
на |
на дистальном |
|||
|
|
|
|
|
||
|
|
|
|
|
проксимальном |
|
|
|
|
|
|
конце |
|
|
|
|
|
|
конце |
|
|
|
|
|
|
|
|
|
|
|
||||
Рефлекторное раздражение нерва с пе- |
|
4 8, —8 |
||||
ререзанным дистальным |
концом |
|
|
|||
Электрическое раздражение нерва с пе- |
19, 40, 37 |
—5,-7,0,2 |
||||
ререзанным дистальным и прокси- |
|
|||||
мальным концами |
|
|
|
|
||
Электрическое |
раздражение |
нерва |
36, 41, 21 |
2, 5, —5 |
||
с лигатурами на дистальном или про- |
|
|||||
ксимальном концах |
|
|
|
|
||
Проведение |
вторичного |
излучения |
44,23, 46,32 |
10, 2, —12,5 |
||
нерва |
с лигатурами на |
проксималь- |
|
|||
ном или дистальном концах |
|
|
|
|||
Проведение |
вторичного |
излучения |
31,31,32,54 |
5, —5, 8 |
||
нерва |
при |
обжигании |
проксималь- |
|
||
ного |
или дистального концов |
|
|
|
||
|
|
|
|
|
|
|
Таким образом, воздействие со стороны мышц является необходимым условием для излучения нерва при искусственном раздражении Ясно выраженная обратная зависимость наблюдается на чисто чувствительных
127

нервах, способность к излучению которых при раздражении определяется связью нерва с центрами.
Излучение спинных кожных нервов (на целом животном) при электризации нерва составляло:
|
|
|
|
% |
В |
норме |
|
|
39; 42; 30; 50; |
|
|
|
|
30; |
После перерезки проксималь- |
0;1Г,—8;2;2; |
|||
|
|
ного |
конца |
6; 0; 4; —2; |
» |
» |
дистального |
|
|
|
|
конца |
. . 30; 35; 25; 28. |
|
Дальнейшие опыты, в которых применялось рецепторное, т. е. физиологическое, раздражение чувствительного нерва расширили наше представление о степени и характере взаимодействия центров и периферии.
Путем раздражения кожных рецепторов возбуждался задний кожный бедренный нерв лягушки, входящий, как известно, в ствол седалищного нерва. При этом ставился следующий вопрос: отразится ли на излучении n. Cutaneus fem. posterior перерезка дистального конца седалищного нерва? Отличие такой постановки опыта от предыдущих состояло в том, что кожная веточка непосредственно не травмировалась. Можно было, однако, предполагать, что изменения состояния нервных центров вследствие перерезки седалищного нерва (т. е. нарушения его связи с большой группой мышц всей нижней части лапки и с кожными рецепторами в этой области) должны будут отразиться на состоянии, а следовательно, и на излучении п. Cutaneus.
На декапитированных, а в некоторых случаях и на целых лягушках осторожно отпрепаровывался на внутренней поверхности кожи бедра участок п. Cutaneus posterior и испытывалось его излучение как в нормальном состоянии всего нервного ствола, так и через некоторое время после перетяжки или перерезки дистального конца седалищного нерва. Опыты производились без применения внешних раздражений.
При тоническом состоянии покоя эффект излучения составлял:
|
% |
До перерезки |
седалищного |
нерва |
22,25,32,33 |
После перерезки |
седалищного |
нерва |
18,21,30,35. |
128
Таким образом, изменение состояния функциональной системы не отражается на общей интенсивности излучения.
Другие результаты наблюдаются, однако, при возбуждении кожной ветки (табл. 21).
ТАБЛИЦА 21
Излучение п. Cutaneus femoris posterior при различных раздражениях кожных рецепторов
|
|
Излучение в % |
|
Характер раздражений |
|
|
|
|
|
до перерезки нерва |
после перерезки |
|
|
|
|
Аппликация 0,2N H2SO |
4 |
25; 22; 24; 40 |
8; 8%; —2; - 6 |
Груз 20 г |
|
35; 45 |
—2; 13 |
Пощипывание кожи |
|
33; 24; 30; 46 |
3; 5; 9; 4 |
|
|
|
|
Интенсивность излучения, связанного с возбужденными состояниями, резко понижается после перерезки седалищного нерва. Мы стоим здесь, несомненно, перед сложной задачей, к которой будем еще возвращаться в связи с дальнейшим анализом процессов взаимодействия.
Ясность в этот вопрос должен внести спектральный анализ, и изучение спектра при этих условиях является поэтому очень важным.
Пока мы можем дать только общую формулировку. Для возникновения излучения при физиологическом возбуждении нерза требуется больше условий, чем при состоянии покоя. При возбуждении необходимой является не только связь нерва и с центральными элементами, и с периферией, но и полноценное нормальное состояние всей системы в целом. Но полученные результаты подчеркивают и другую сторону вопроса, которая будет рассмотрена дальше, — значительно большую диффузность возникающих в центральной нервной системе состояний, чем это можно было предполагать раньше. Ведь исчезновение излучения п. Cutaneus показывает, что, несмотря на то, что данная рефлекторная дуга не была непосредственно затронута, какое-то изменение состояния захватило и соответствующие ей центры.
9 А. А. Гурвнч |
129 |