

14.Пути образования и превращения пвк
Пировиноградная кислота — химическое соединение с формулой СН3СОСООН, органическая кетокислота. Пируваты (соли пировиноградной кислоты) — важные химические соединения в биохимии. Они являются конечным продуктом метаболизма глюкозы в процессе гликолиза. Одна молекула глюкозы превращается при этом в две молекулы пировиноградной кислоты. Дальнейший метаболизм пировиноградной кислоты возможен двумя путями — аэробным и анаэробным.
В условиях достаточного поступления кислорода пировиноградная кислота превращается в ацетил-кофермент А, являющийся основным субстратом для серии реакций, известных как цикл Кребса, или дыхательный цикл, цикл трикарбоновых кислот. Пируват также может быть превращён в анаплеротической реакции в оксалоацетат. Оксалоацетат затем окисляется до углекислого газа и воды. Если кислорода недостаточно, пировиноградная кислота подвергается анаэробному расщеплению с образованием молочной кислоты у животных и этанола у растений. При анаэробном дыхании в клетках пируват, полученный при гликолизе, преобразуется в лактат при помощи фермента лактатдегидрогеназы и NADP в процессе лактатной ферментации, либо в ацетальдегид и затем в этанол в процессе алкогольной ферментации». Пировиноградная кислота является «точкой пересечения» многих метаболических путей.
Пути образования и превращения: 1.Удаление молочной кислоты, накопившейся при анаэробном дыхании. Мол. Кис-та поступает в кровь и переносится из мышц в печень, где она окисляется с образованием ПВК и восстановительного НАД. 2. Гликолиз(схоже со спиртовым брожением). Вначале глюкоза подвергается фосфорилированию с образованием гексозофосфорных эфиров, расхождение начинается лишь на стадии образования ПВК(роль акцептора водорода, отцепляемого от НАД) 3. Разложение жиров в анаэробных условиях. Глицерин через стадию образования ПВК сбраживается с образованием различных продуктов. 4. Расщепление цистеина. Под влиянием десульфогидрогеназы приводит к образованию ПВК и сероводорода.
15.Синтез жира из углеводов.
Процесс синтеза жирных кислот длиннее, чем их окисление
Биосинтез жирных кислот наиболее активно происходит в цитозоле клеток печени, кишечника, жировой ткани в состоянии покоя или после еды.
Условно можно выделить 4 этапа биосинтеза:
1. Образование ацетил-SКоА из глюкозы, других моносахаров или кетогенных аминокислот.
2. Перенос ацетил-SКоА из митохондрий в цитозоль:
может быть в комплексе с карнитином, подобно тому как переносятся внутрь митохондрии высшие жирные кислоты, но здесь транспорт идет в другом направлении,
обычно в составе лимонной кислоты, образующейся в первой реакции ЦТК.
Поступающий из митохондрий цитрат в цитозоле расщепляется АТФ-цитрат-лиазой до оксалоацетата и ацетил-SКоА.
Оксалоацетат в дальнейшем восстанавливается до малата, и последний либо переходит в митохондрии (малат-аспартатный челнок), либо декарбоксилируется в пируват малик-ферментом ("яблочный" фермент).
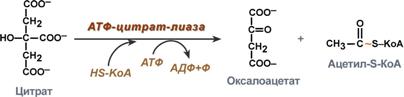
Образование ацетил-SКоА из лимонной кислоты
3. Образование малонил-SКоА из ацетил-SКоА.
Карбоксилирование ацетил-SКоА катализируется ацетил-SКоА-карбоксилазой, мульферментным комплексом из трех ферментов.

Образование малонил-SКоА из ацетил-SКоА
4. Синтез пальмитиновой кислоты.
Осуществляется мультиферментным комплексом "синтаза жирных кислот" (синоним пальмитатсинтаза) в состав которого входит 6 ферментов и ацил-переносящий белок (АПБ).
Ацил-переносящий белок включает производное пантотеновой кислоты – 6-фосфопантетеин (ФП), имеющий HS-группу, подобно HS-КоА. Один их ферментов комплекса, 3-кетоацил-синтаза, также имеет HS-группу в составе цистеина. Взаимодействие этих групп обусловливает начало и продолжение биосинтеза жирной кислоты, а именно пальмитиновой кислоты. Для реакций синтеза необходим НАДФН.
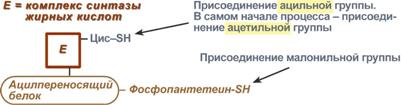
Активные группы синтазы жирных кислот
В первых двух реакциях последовательно присоединяются малонил-SКоА к фосфопантетеину ацил-переносящего белка и ацетил-SКоА к цистеину 3-кетоацилсинтазы.
3-Кетоацилсинтаза катализирует третью реакцию – перенос ацетильной группы на С2малонила с отщеплением карбоксильной группы.
Далее кетогруппа в реакциях восстановления (3-кетоацил-редуктаза), дегидратации (дегидратаза) и опять восстановления (еноил-редуктаза) превращается в метиленовую с образованием насыщенного ацила, связанного с фосфопантетеином.
Ацилтрансфераза переносит полученный ацил на цистеин 3-кетоацил-синтазы, к фосфопантетеину присоединяется малонил-SКоА и цикл повторяется 7 раз до образования остатка пальмитиновой кислоты. После этого пальмитиновая кислота отщепляется шестым ферментом комплекса тиоэстеразой.