

Стволовые центры
Центры, находящиеся в стволе мозга (продолговатый мозг, мост, средний мозг), управляют висцеральными функциями посредством парасимпатических волокон, проходящих в составе блуждающего, языкоглоточного, лицевого и глазодвигательного нервов.
В продолговатом мозгу находятся нервные центры, с помощью которых осуществляются сложные рефлексы, такие как жевание, глотание, слюноотделение.
Здесь же расположены центры, тормозящие сердечную деятельность, и стимулирующие секрецию желудочных желез.
Рефлекторные процессы, происходящие в ядерных образованиях спинного продолговатого, среднего мозга и моста, находятся под постоянным влиянием гипоталамуса — высшего центра регуляции висцеральных функций.
Гипоталамические центры
Гипоталамус управляет всеми основными гомеостатическими функциями организма. Интегративная функция гипоталамуса обеспечивается автономными, соматическими и эндокринными механизмами. Он представляет собой скопление 32 пар ядер, которые условно разделяются на три группы: передние, средние и задние.
Эфферентные пути гипоталамуса связывают его с ретикулярной формацией ствола мозга, ядрами спинного мозга.
Нисходящие влияния гипоталамуса обеспечивают регуляцию функций через автономную нервную систему.
Важным компонентом в осуществлении нисходящих влияний гипоталамуса являются и гормоны, гипофиза.
Лимбическая система
Лимбическая система это совокупность функционально связанных между собой образований древней коры (гиппокамп) , старой коры (поясная извилина) и подкорковых структур (миндалевидный комплекс, ряд ядер таламуса и гипоталамуса.
Наряду с управлением висцеральными функциями лимбическая система участвует в эмоциональном и инстинктивном (пищевом, половом, оборонительном) поведении.
Она оказывает также влияние на смену фаз сна и бодрствования.
Общие свойства нервных центров
Нервные центры представляют собой совокупность нейронов, расположенных в одной или нескольких структурах ЦНС и регулирующих определенную функцию организма.
Классификация центров:
Классификация по расположению
Классификация по функциональной роли
Принцип работы центров –рефлекторный.
Афферентный вход Центры суммируют входящую информацию
Нейронный ансамбль Информация видоизменяется благодаря взаимодействию нейронов
Эфферентная часть. Несколько мест
Общие принципы деятельности нервных центров.
Автоматия центров (спонтанная активность, обусловлена метаболизмом).
Тонус центров (обусловлен потоком импульсов от различных рецептивных полей, действием на нейроны биологически активных веществ и метаболитов).
Пластичность центров
Пластичность - способность объединяться в разные ансамбли и связываться с разными центрами.
Пластичность обусловлена
1) способностью синапсов к тренировке (кальций, медиаторы, рецепторы),
2) морфологическими изменениями – синтез новых мембранных рецепторов, рост шипиков на дендритах.
3) Способность к временному доминированию.
Вегетативный индекс Кердо
Чаще всего это понятие используется в спортивной медицине для оценки функциональных возможностей, тренированности организма испытуемого. Тем не менее индекс Кердо изначально предусматривался в рамках анализа деятельности вегетативной нервной системы.
Рассчитать индекс Кердо несложно. Для этого нужны показатели пульса и диастолического артериального давления. Рассчитывается отношение частоты сердечных сокращений к показателю ДАД в виде дроби. От единицы отнимают получившееся значение и умножают для удобства расчетов на 100.

Понятия нормы индекса Кердо не существует. Этот показатель говорит о преобладании тонуса парасимпатической части вегетативной нервной системы при отрицательном значении. При индексе Кердо больше нуля речь ведется об обратном состоянии – симпатикотонии, когда вегетативный тонус смещен в сторону преобладания симпатической его части.
Принципы гормональной регуляции: прямая и обратная регуляторная связь.
Специфические регуляторы, которые секретируются эндокринными железами в кровь или лимфу, а затем попадают на клетки-мишени, называют гормонами.
Прямая
Функциональная активность эндокринной железы может регулироваться «субстратом», на который направлено действие гормона. Так, глюкоза стимулирует секрецию инсулина из β-клеток панкреатических островков (островки Лангерганса), а инсулин понижает концентрацию глюкозы в крови, активируя ее транспорт в мышцы и печень. Это происходит следующим образом. Глюкоза входит в β-клетки поджелудочной железы через переносчик глюкозы и сразу же фосфорилируется глюкокиназой, после чего вовлекается в гликолиз. Образующийся при этом АТФ ингибирует калиевые каналы, вследствие чего снижается мембранный потенциал β-клеток и активируются потенциалзависимые кальциевые каналы. Входящий в β-клетку кальций стимулирует слияние везикул, содержащих инсулин, с плазматической мембраной и секрецию инсулина. Инсулин активирует перенос глюкозы в печень, сердце и скелетные мышцы, вследствие чего уровень глюкозы в крови снижается, замедляется ее вход в β-клетки и уменьшается секреция инсулина (рис. 5.1).
Такой же механизм лежит в основе секреции паратгормона (паратиреоидный гормон, паратирин) и кальцитонина. Оба гормона влияют на концентрацию кальция и фосфатов в крови. Паратиреоидный гормон вызывает выход минеральных веществ из кости и стимулирует реабсорбцию кальция в почках и кишечнике, в результате чего возрастает концентрация кальция в плазме крови. Кальцитонин, напротив, стимулирует поступление кальция и фосфатов в костную ткань, в результате чего концентрация минеральных веществ в крови снижается. При высокой концентрации кальция в крови подавляется секреция паратиреоидного гормона и стимулируется секреция кальцитонина. В случае снижения концентрации кальция в крови секреция паратиреоидного гормона усиливается, а кальцитонина — ослабляется.
Такая регуляция постоянства внутренней среды организма, происходящая по принципу отрицательной обратной связи, очень эффективна для поддержания гомеостаза, однако не может выполнять все задачи адаптации организма. Например, кора надпочечников продуцирует стероидные гормоны в ответ на голод, болезнь, эмоциональное возбуждение и т. п. Чтобы эндокринная система могла «отвечать» на свет, звуки, запахи, эмоции и т. д., должна существовать связь между эндокринными железами и нервной системой.
Основные связи между нервной и эндокринной системами регуляции осуществляются посредством взаимодействия гипоталамуса и гипофиза. Нервные импульсы, приходящие в гипоталамус, активируют секрецию так называемых рилизинг-факторов (либеринов и статинов): тиреолиберина, соматолиберина, пролактолиберина, гонадолиберина и кортиколиберина, а также соматостатина и пролактостатина. Мишенью для либеринов и статинов, секретируемых гипоталамусом, является гипофиз. Каждый из либеринов взаимодействует с определенной популяцией клеток гипофиза и вызывает в них синтез соответствующих тропинов: тиреотропина, соматотропного гормона (соматотропин — гормон роста), пролактина, гонадотропного гормона, (гонадотропины — лютеинизирующий и фолликулостимулирующий), а также адренокортикотропного гормона (АКТГ, кортикотропин). Статины оказывают на гипофиз влияние, противоположное действию либеринов, — подавляют секрецию тропинов. Тропины, секретируемые гипофизом, поступают в общий кровоток и, попадая на соответствующие железы, активируют в них секреторные процессы.
Обратная
Регуляция деятельности гипофиза и гипоталамуса, кроме сигналов, идущих «сверху вниз», осуществляется гормонами «исполнительных» желез. Эти «обратные» сигналы поступают в гипоталамус и затем передаются в гипофиз, что приводит к изменению секреции соответствующих тропинов.
Обратные связи не только позволяют регулировать концентрацию гормонов в крови, но и участвуют в дифференцировке гипоталамуса в онтогенезе. Образование половых гормонов в женском организме происходит циклически, что объясняется циклической секрецией гонадотропных гормонов. Синтез этих гормонов контролируется гипоталамусом, образующим рилизинг-фактор этих тропинов (гонадолиберин).
В железах внутренней секреции иннервированы, как правило, только сосуды, а эндокринные клетки изменяют свою биосинтетическую и секреторную активность лишь под действием метаболитов, кофакторов и гормонов, причем не только гипофизарных. Так, ангиотензин II стимулирует синтез и секрецию альдостерона. Отметим также, что некоторые гормоны гипоталамуса и гипофиза могут образовываться не только в этих тканях. Например, соматостатин (гормон гипоталамуса, ингибирующий образование и секрецию гормона роста) обнаружен также в поджелудочной железе, где он подавляет секрецию инсулина и глюкагона.
Большинство нервных и гуморальных путей регуляции сходится на уровне гипоталамуса и благодаря этому в организме образуется единая нейроэндокринная регуляторная система. К клеткам гипоталамуса подходят аксоны нейронов, расположенных в коре больших полушарий и подкорковых образованиях. Эти аксоны секретируют различные нейромедиаторы, оказывающие на секреторную активность гипоталамуса как активирующее, так и тормозное влияние. Поступающие из мозга нервные импульсы гипоталамус «превращает» в эндокринные стимулы, которые могут быть усилены или ослаблены в зависимости от гуморальных сигналов, поступающих в гипоталамус от желез и тканей, подчиненных ему.
Тропины, образующиеся в гипофизе, не только регулируют деятельность подчиненных желез, но и выполняют самостоятельные эндокринные функции. Например, пролактин оказывает лактогенное действие, а также тормозит процессы дифференцировки клеток, повышает чувствительность половых желез к гонадотропинам, стимулирует родительский инстинкт. Кортикотропин является не только стимулятором стероцдогенеза, но и активатором липолиза в жировой ткани, а также важнейшим участником процесса превращения в мозге кратковременной памяти в долговременную. Гормон роста может стимулировать активность иммунной системы, обмен липидов, Сахаров и т. д.
В задней доле гипофиза (нейрогипофиз) депонируются антидиуретический гормон (вазопрессин) и окситоцин (см. рис. 5.3). Первый вызывает задержку воды в организме и повышает тонус сосудов, второй стимулирует сокращение матки при родах и секрецию молока. Оба гормона синтезируются в гипоталамусе, затем транспортируются по аксонам в заднюю долю гипофиза, где депонируются и потом секретируются в кровь.
Характер процессов, протекающих в ЦНС, во многом определяется состоянием эндокринной регуляции. Так, андрогены и эстрогены формируют половой инстинкт, многие поведенческие реакции. Очевидно, что нейроны, точно так же как и другие клетки нашего организма, находятся под контролем гуморальной системы регуляции. Нервная система, эволюционно более поздняя, имеет как управляющие, так и подчиненные связи с эндокринной системой. Эти две регуляторные системы дополняют друг друга, образуют функционально единый механизм, что обеспечивает высокую эффективность нейрогуморальной регуляции, ставит ее во главе систем, согласующих все процессы жизнедеятельности в многоклеточном организме.
Особенности биосинтеза, секреции и транспорта гормонов разной химической природы.
Белково-пептидные гормоны. В процессе образования белковых и пептидных гормонов в клетках эндокринных желез происходит образование полипептида, не обладающего гормональной активностью. Но такая молекула в своем составе имеет фрагмент(ы), содержащий(е) аминокислотную последовательность данного гормона. Такая белковая молекула называется пре-про-гормоном и имеет в своем составе (обычно на N-конце) структуру, которая называется лидерной или сигнальной последовательностью (пре-). Эта структура представлена гидрофобными радикалами и нужна для прохождения этой молекулы от рибосом через липидные слои мембран внутрь цистерн эндоплазматического ретикулума (ЭПР). При этом, во время перехода молекулы через мембрану в результате ограниченного протеолиза лидерная (пре-) последовательность отщепляется и внутри ЭПР оказывается прогормон. Затем через систему ЭПР прогормон транспортируется в комплекс Гольджи и здесь заканчивается созревание гормона. Вновь в результате гидролиза под действием специфических протеиназ отщепляется оставшийся (N-концевой) фрагмент (про-участок). Образованная молекула гормона, обладающая специфической биологической активностью поступает в секреторные пузырьки и накапливается до момента секреции.
При синтезе гормонов из числа сложных белков гликопротеинов (например, фолликулостимулирующего (ФСГ) или тиреотропного (ТТГ) гормонов гипофиза) в процессе созревания происходит включение углеводного компонента в структуру гормона.
Производные аминокислот. Из тирозина синтезируются гормоны мозгового слоя надпочечников адреналин и норадреналин, а также йодсодержащие гормоны щитовидной железы. В ходе синтеза адреналина и норадреналина тирозин подвергается гидроксилированию, декарбоксилированию и метилированию с участием активной формы аминокислоты метионина.
В щитовидной железе происходит синтез йодсодержащих гормонов трийодтиронина и тироксина (тетрайодтиронина). В ходе синтеза происходит йодирование фенольной группы тирозина. Из остатков тирозина образуются моно- и ди-иодтирозин. Из них примерно 30 % остатков в результате конденсации могут превратитьться в три- и тетра- иодтиронины. Конденсация и иодирование идут с участием одного и того же фермента — тиреопероксидазы. Дальнейшее созревание гормонов щитовидной железы происходит в железистых клетках — ТГ поглощается клетками путем эндоцитоза и образуется вторичная лизосома в результате слияния лизосомы с поглощенным белком ТГ.
Протеолитические ферменты лизосом обеспечивают гидролиз ТГ и образование Т3 и Т4, которые выделяются во внеклеточное пространство. А моно- и дииодтирозин деиодируются с помощью специального фермента деиодиназы и иод повторно может подвергаться органификации. Для синтеза тиреоидных гормонов характерным является механизм торможения секреции по типу отрицательной обратной связи (Т3 и Т4 угнетают выделение ТТГ).
Стероидные гормоны. В эту группу входят тестостерон, эстрадиол, эстрон, прогестерон, кортизол, альдостерон и др. Эти гормоны образуются из холестерина в корковом веществе надпочечников (кортикостероиды), а также в семенниках и яичниках (половые стероиды). Свободный холестерин поступает в митохондрии, где превращается в прегненолон, который затем попадает в эндоплазматическую сеть и после этого — в цитоплазму.
Особенности секреции и транспорта гормонов разной химической природы.
Секреция
Процессы секреции гормонов тесно сопряжены с процессами их биосинтеза. Степень их сопряженности зависит от химической природы гормона и особенностями механизмов его секреции. Подразделяют:
Освобождение гормонов из клеточных секреторных гранул (секреция белково-пептидных гормонов и катехоламинов )
Освобождение гормонов из белковосвязанной формы ( секреция тиреоидных гормонов )
Относительно свободная диффузия гормонов через клеточные мембраны ( стероидные гормоны )
Транспорт
Гормоны, имеющие гидрофильную природу (катехоламины, серотонин, белково-пептидные и др.), синтезируются «впрок» и выделяются в кровь определенными порциями за счет опустошения секреторных везикул. Уровень этих гормонов в крови возрастает при увеличении частоты выброса гормона из клеток эндокринной железы. В отличие от этого стероидные и тиреоидине гормоны, а также эйкозаноиды не накапливаются в специальных структурах клетки, а благодаря своей липофильности свободно проходят через плазматическую мембрану эндокринной клетки и попадают в кровь. Содержание этих гормонов в крови регулируется ускорением или замедлением их синтеза.
Поступая в кровь, гормоны связываются с белками плазмы. Обычно лишь 5—10% молекул гормонов находится в крови в свободном состоянии, и только они могут взаимодействовать с рецепторами. К числу специфических гормонсвязывающих белков относятся транскортин, связывающий кортикостероиды, тестостерон-эстрогенсвязывающий глобулин, тироксинсвязывающий глобулин и т. д. Альдостерон, по-видимому, не имеет специфических «транспортных» белков, поэтому находится преимущественно в связи с альбумином
Виды и пути действия гормонов на клетки-мишени.
Различают пять видов действия гормонов на ткани-мишени: метаболическое, мор-фогенетическое, кинетическое, корригирующее и реактогенное.
Метаболическое действие гормонов — вызывает изменение обмена веществ в тканях. Оно происходит за счет трех основных гормональных влияний. Во-первых, гормоны меняют проницаемость мембран клетки и органоидов, что изменяет условия мембранного транспорта субстратов, ферментов, ионов и метаболитов и, соответственно, все виды метаболизма. Во-вторых, гормоны меняют активность ферментов в клетке, приводя к изменению их структуры и конфигурации, облегчая связи с кофакторами, уменьшая или увеличивая интенсивность распада ферментных молекул, стимулируя или подавляя активацию проферментов. В-третьих, гормоны изменяют синтез ферментов, индуцируя или подавляя их образование за счет влияния на генетический аппарат ядра клетки, как прямо вмешиваясь в процессы синтеза нуклеиновых кислот и белка, так и опосредованно через энергетическое и субстратно- ферментное обеспечение этих процессов. Сдвиги метаболизма, вызываемые гормонами, лежат в основе изменения функции клеток, ткани или органа.
Морфогенетическое действие — влияние гормонов на процессы формообразования, дифференцировки и роста структурных элементов. Осуществляются эти процессы за счет изменений генетического аппарата клетки и обмена веществ. Примерами может служить влияние соматотропина на рост тела и внутренних органов, половых гормонов — на развитие вторичных половых признаков.
Кинетическое действие — способность гормонов запускать деятельность эффектора, включать реализацию определенной функции. Например, окситоцин вызывает сокращение мускулатуры матки, адреналин запускает распад гликогена в печени и выход глюкозы в кровь, вазопрессин включает обратное всасывание воды в собирательных трубочках нефрона, без него не происходящее.
Корригирующее действие — изменение деятельности органов или процессов, которые происходят и в отсутствие гормона. Примером корригирующего действия гормонов является влияние адреналина на частоту сердечных сокращений, активация окислительных процессов тироксином, уменьшение обратного всасывания ионов калия в почках под влиянием альдостерона. Разновидностью корригирующего действия является нормализующий эффект гормонов, когда их влияние направлено на восстановление измененного или даже нарушенного процесса. Например, при исходном превалировании анаболических процессов белкового обмена глюкокортикоиды вызывают катаболический эффект, но если исходно преобладает распад белков, глюкокортикоиды стимулируют их синтез.
В более широком плане зависимость величины и направленности эффекта гормона от имеющихся перед его действием особенностей метаболизма или функции определяется правилом исходного состояния, описанном в начале главы. Правило исходного состояния показывает, что гормональный эффект зависит не только от количества и свойств молекул гормона, но и от реактивности эффектора, определяемой числом и свойствами мембранных рецепторов к гормону. Реактивностью в рассматриваемом контексте называют способность эффектора реагировать определенной величиной и направленностью ответа на действие конкретного химического регулятора.
Реактогенное действие гормонов — способность гормона менять реактивность ткани к действию того же гормона, других гормонов или медиаторов нервных импульсов. Так, например, кальцийрегули-рующие гормоны снижают чувствительность дистальных отделов нефрона к действию вазопрессина, фолликулин усиливает действие прогестерона на слизистую оболочку матки, тиреоидные гормоны усиливают эффекты катехоламинов. Разновидностью реактогенного действия гормонов являетсяпермиссивное действие, означающее способность одного гормона давать возможность реализоваться эффекту другого гормона. Так, например, глюкокортикоиды обладают пермиссивным действием по отношению к катехоламинам, т.е. для реализации эффектов адреналина необходимо присутствие малых количеств кортизола, инсулин обладает пермиссивным действием для соматотропина (гормона роста) и др. Особенностью гормональной регуляции является то, что реактогенное действие гормоны могут реализовать не только в тканях — мишенях, где концентрация рецепторов к ним высока, но и в других тканях и органах, имеющих единичные рецепторы к гормону.
Пути действия гормонов на клетки-мишени.
Пути действия гормонов рассматриваются в виде двух альтернативных возможностей: 1) действия гормона с поверхности клеточной мембраны после связывания со специфическим мембранным рецептором и запуска тем самым цепочки биохимических превращений в мембране и цитоплазме (эффекты пептидных гормонов и катехола-минов); 2) действия гормона путем проникновения через мембрану и связывания с рецептором цитоплазмы, после чего гормон-рецеп-торный комплекс проникает в ядро и органоиды клетки, где и реализует свой регуляторный эффект (стероидные Гормоны, гормоны щитовидной железы). Считается, что функция распознавания предназначенного определенным клеткам специфического гормонального сигнала у всех клеток для всех гормонов осуществляется мембранным рецептором, а после связывания гормона с соответствующим ему рецептором, дальнейшая роль гормон- рецепторного комплекса для пептидных и стероидных гормонов различна.
Молекулярные механизмы действия гормонов разной химической природы на клетки-мишени.
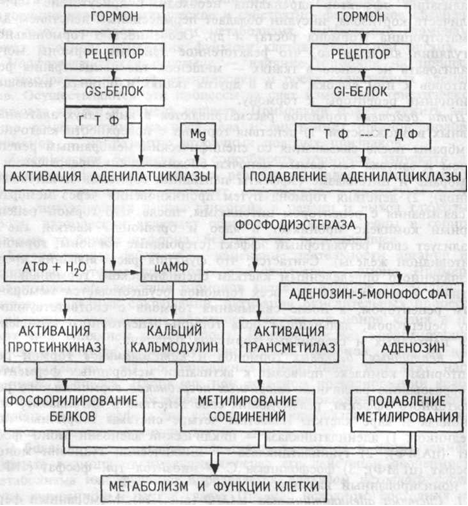
Система «аденилатциклаза – цАМФ»
Мембранный фермент аденилатциклаза может находиться в двух формах — активированной и неактивированной. Активация аденилатциклазы происходит под влиянием гормон-рецепторного комплекса, образование которого приводит к связыванию гуанилового нуклеотида (ГТФ) с особым регуляторным стимулирующим белком (GS-белок), после чего GS-белок вызывает присоединение Mg к аденилатциклазе и ее активацию. ак действуют активирующие аденилатциклазу гормоны — глюкагон, тиротропин, паратирин, вазопрессин (через V-2-рецепторы), гонадотропин и др. Ряд гормонов, напротив, подавляет аденилатциклазу — соматостатин, ангиотензин- II и др. Гормон-рецепторные комплексы этих гормонов взаимодействуют в мембране клетки с другим регуляторным ингибирующим белком (GI-белок), который вызывает гидролиз ГТФ до ГДФ и, соответственно, подавление активности аденилатциклазы. Адреналин через бета-адренорецепторы активирует аденилатциклазу, а через альфа-1-адренорецепторы ее подавляет.
Под влиянием аденилатциклазы из АТФ синтезируется цАМФ, вызывающий активацию двух типов протеинкиназ в цитоплазме клетки, ведущих к фосфорилированию многочисленных внутриклеточных белков. Это меняет проницаемость мембран, активность и количество ферментов, т.е. вызывает типичные для гормона метаболические и, соответственно, функциональные сдвиги. В табл.3.3 приведены основные эффекты активации цАМФ-зависимых протеинкиназ. Секреторный, двигательный эффекты или изменение конфигурации клетки
Кроме активации протеинкиназ внутриклеточные эффекты цАМФ реализуются также через три других механизма:
1) Систему кальций-кальмодулин,
2) Трансметилазную систему,
3) Аденозин-5-монофосфат — аденозин.
О роли системы кальций-кальмодулин будет сказано ниже. Трансметилазная система обеспечивает метилирование ДНК, всех типов РНК, белков хроматина и мембран, ряда гормонов на уровне тканей, фосфолипидов мембран. Это способствует реализации многих гормональных влияний на процессы пролиферации, дифференцировки, состояние проницаемости мембран и свойства их ионных каналов и, что важно подчеркнуть особо, влияет на доступность мембранных реиепторных белков молекулам гормонов.
Прекращение гормонального эффекта, реализуемого через систему аденилатциклаза-цАМФ, осуществляется с помощью специального фермента фосфодиэстеразы цАМФ, вызывающей гидролиз этого вторичного посредника с образованием аденозин-5-монофосфата. Однако, этот продукт гидролиза превращается в клетке в аденозин, обладающий четким эффектом подавления процессов метилирования, что также играет роль в реализации определенных гормональных влияний.
Система гуанилатциклаза-цГМФ.
Активация мембранной гу-анилатциклазы происходит не под непосредственным влиянием гормон-рецепторного комплекса, а опосредованно через ионизированный кальций и оксидантные системы мембран. Типичная стимуляция активности гуанилатциклазы ацетилхолином также реализуется опосредованно через Са++ . Через активацию гуанилатциклазы реализует эффект и натриуретический гормон предсердий — атриопептид. Путем активации перекисного окисления стимулирует гу-анилатциклазу биологически активное вещество (тканевой гормон) сосудистой стенки — расслабляющий эндотелиальный фактор. Под влиянием гуанилатциклазы из ГТФ синтезируется цГМФ, активирующий цГМФ-зависимые протеинкиназы, которые уменьшают скорость фосфорилирования легких цепей миозина в гладких мышцах стенок сосудов, приводя к их расслаблению. В большинстве тканей биохимические и физиологические эффекты цАМФ и цГМФ проти-воложны. Примерами могут служить стимуляция сокращений сердца под влиянием цАМФ и торможение их цГМФ, стимуляция сокращения гладких мышц кишечника цГМФ и подавление цАМФ. цГМФ играет роль в гиперполяризации рецепторов сетчатки глаза под влиянием фотонов света. Ферментативный гидролиз цГМФ осуществляется с помощью специфической фосфодиэстеразы.2. Система «гуанилатциклаза – цГМФ»
Системы «фосфолипаза С – ИТФ» и «Са – кальмодулин».
Гормонрецепторный комплекс с участием регуляторного G-белка ведет к активации мембранного фермента фосфолипазы С, вызывающей гидролиз фосфолигшдов мембраны с образованием двух вторичных посредников: инозитол-3-фосфата и диадилглицерола. Инозитол-3-фосфат вызывает выход Са++ из внутриклеточных депо, в основном из эндоплазматического ретикулума, ионизированный кальций связывается со специализированным белком кальмодулином, что обеспечивает активацию ряда протеинкиназ и фосфорилирование внутриклеточных структурных белков и ферментов
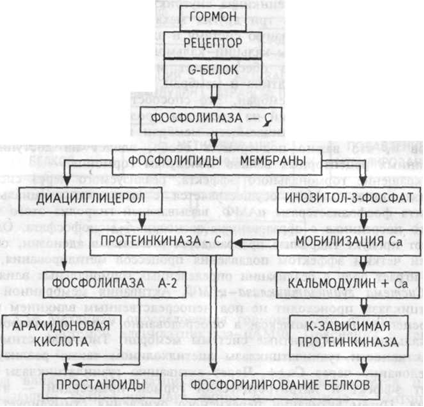
Механизм опосредования гормонального стимула системой фосфолипаза С — инозитол-3-фосфат. Пояснения в тексте.
В свою очередь диацилглицерол способствует резкому повышению сродства протеинкиназы С к ионизированному кальцию, последний без участия кальмодулина ее активирует, что также завершается процессами фосфорилирования других белков. Диацилглицерол одновременно может реализовывать и другой путь опосредования гормонального эффекта, так как он активирует фосфолипазу А-2. Под влиянием последней из мембранных фосфолипидов образуется арахидоновая кислота, являющаяся источником мощных по метаболическим и физиологическим эффектам веществ — простагландинов и лейкотриенов. Необходимо указать, что в разных клетках превалирует один или другой пути образования вторичных посредников, что в конечном счете и определяет физиологический эффект гормона. Через рассмотренные системы вторичных посредников реализуются эффекты адреналина (при связи с альфа-адренорецептором), вазопрессина (при связи с V- 1-рецептором), ангиотензина- II, соматостатина, окситоцина и других гормонов.
Система «фосфолипаза А2 – ДАГ».
гормон-рецептор - G-белок - фосфолипаза А2 - ДАГ - киназа С - фосфорилирование белка - физиологический ответ.
Нейросекреторная функция гипоталамуса. Рилизинг-факторы, их характеристика. Гипоталамо-гипофизарные связи.
Нейросекреторная функция гипоталламуса
Многочисленные исследования показали важную роль нейросекреторных клеток гипоталамуса в регуляции функций внутренних органов. Эта регуляция может осуществляться либо прямым путем, либо посредством желез внутренней секреции.
Особое значение нейросекреторные элементы имеют в осуществлении защитных и приспособительных реакций организма.
В супраоптическом и паравентрикулярном ядрах располагаются клеточные тела, отростки которых образуют гипоталамо-гипофизарный тракт, идущий в заднюю долю гипофиза.
Последняя выступает в роли гемального органа, где наполненные секретом аксонные терминали функционально взаимодействуют с выносящими секрет сосудами. Клетки супраоптического и паравентрикулярных ядер получают синаптический вход от разных структур мозга. Это типичные нейроны.
Они генерируют потенциалы действия, распространяющиеся по аксону. Достигая аксонной терминали, потенциал вызывает секрецию пептидного гормона, которая осуществляется по механизму экзоцитоза, т. е. того самого механизма, с помощью которого выделяются и медиаторы.По своей ультраструктуре эти клетки отличаются от других нейронов наличием крупных (до 190 нм) электронно-плотных секреторных гранул, которые окружены специальной мембраной. Особенно много гранул в аксональных терминалях.
В области секреторной мембраны аксона локализуются мелкие пузырьки, не имеющие плотного содержимого. Они являются, по-видимому, либо продуктами пиноцитоза, либо остатками изливших содержимое секреторных гранул.
Благодаря специальным исследованиям установлено, что клетки супраоптического ядра вырабатывают вазопрессин, паравентрикулярного — окситоцин.
Секретируемый нервными клетками материал перемещается по аксонам гипофизарно-гипоталамического тракта в форме коллоидных гранул. Перерезка тракта сопровождается накоплением нейросекрета в его проксимальном конце и истощением в дистальном. Различного рода стрессорные воздействия, жажда, обезвоживание также ведут к истощению нейросекрета. У крыс, например, стресс вызывает истощение запаса нейросекрета в течение 10 мин.
Не менее важна роль гипоталамуса и в секреции аденогипофизарных гормонов. В состав мелкоклеточных ядер вентральной гипоталамической области входят клетки, которые вырабатывают нейрогормоны, регулирующие секреторную активность тройных желез аденогипофиза.
С ними они связаны посредством специальной портальной системы кровеносных сосудов. Сейчас различается 7 эффектов, которые оказывают эти клетки на железистые образования аденогипофиза. Последнее дает основание говорить о 7 гипоталамических факторах: CRF — фактор, вызывающий выделение кортикотропного гормона; LRF— лютеинизирующего гормона; FSHRF — фолликулостимулирующего гормона; TRF — тиреотропного гормона; GRF или SRF — соматотропного гормона; PIF — фактор, тормозящий выделение пролактина; MIF — тормозящий выделение меланоцитстимулирующего гормона.
Все эти факторы имеют пептидную природу. Следовательно, гипоталамус выполняет роль своеобразного реле, включающего гипофизарную гормональную цепь в регуляцию различных висцеральных
Рилизинг-факторы, их характеристика
Риллизинг-факторы-класс пептидных гормонов гипоталламуса, общим свойством которых является реализафия их эффектовчерез стимуляцию синтеза и секреции в кровь тех или иных тропных гормонов передней доли гипофиза. Подразделяются на 2 группы:
1)Либерины- гормоны белковой природы. Вырабатываются в гипоталамусе, далее поступают в гипофиз и активируют синтез и выделение в кровь гормонов передней доли гипофиза. Основными либеринами являются соматолиберины, кортиколиберины, тиреолиберины и т.д.
2) Статины- гормоны белковой природы. Вырабатываются в гипоталамусе, далее поступают в гипофиз и подавляют синтез и выделение в кровь гормонов передней доли гипофиза. Основными статинами являются соматостатины, кортикостатины, тиреостатины и т.д.
Гипоталамо-гипофизарные связи. Гормоны гипофиза.
Гипоталамо-гипофизарная система — объединение структур гипофиза и гипоталамуса, выполняющее функции, как нервной системы, так и эндокринной. Этот нейроэндокринный комплекс является примером того, насколько тесно связаны в организме млекопитающих нервный и гуморальный способы регуляции.
Строение:Гипоталамо-гипофизарная система состоит из ножки гипофиза, начинающейся в вентромедиальной области гипоталамуса, и трёх долей гипофиза: аденогипофиз (передняя доля), нейрогипофиз (задняя доля) и вставочная доля гипофиза. Работа всех трёх долей управляется гипоталамусом с помощью особых нейросекреторных клеток. Эти клетки выделяют специальные гормоны — рилизинг-гормоны. Релизинг-факторы попадают в гипофиз, а точнее в аденогипофиз через воротную вену гипофиза.
Гормоны гипоталамо-гипофизарной системы:
Под влиянием того или иного типа воздействия гипоталамуса, доли гипофиза выделяют различные гормоны, управляющие работой почти всей эндокринной системы человека. Исключение составляет поджелудочная железа и мозговая часть надпочечников. У них есть своя собственная система регуляции.
Гормоны нейрогипофиза, их функции.
Задняя доля гипофиза (нейрогипофиз)
Гормонами нейрогипофиза являются антидиуретический гормон (АДГ) или вазопрессин и окситоцин. Образование гормонов задней доли гипофиза происходит в основном в ядрах гипоталамуса в результате процессов нейросекреции. АДГ, по-видимому, секретируется в супраоптическом ядре, а окситоцин - в паравентрикулярном ядре гипоталамуса. По аксонам нервных клеток эти гормоны поступают в заднюю долю гипофиза. Этому способствует наличие прямой нервной связи нейрогипофиза с ядрами гипоталамуса.
Механизм действия АДГ состоит в том, что под его воздействием усиливается реабсорбция воды в собирательных трубках почек. При снижении секреции АДГ возникает несахарный диабет, который сопровождается выделением больших количеств мочи (иногда десятки литров в сутки), что связано с нарушением реабсорбции воды в собирательных трубках. В больших дозах он вызывает сокращение гладких мышц сосудов (особенно артериол), что приводит к повышению артериального давления.
Окситоцин стимулирует сокращение гладких мышц матки и, таким образом, обеспечивает нормальное течение родового акта, влияет на отделение молока.
Гормоны аденогипофиза, их функции.
Гормоны передней доли гипофиза(аденогипофиз)
Соматотропин Гормон роста Обладает анаболическим воздействием, следовательно, как любой анаболик, СТ усиливает процессы синтеза (в особенности — белкового). Поэтому соматотропин называют часто «гормоном роста».
При нарушении секреции соматотропина возникает три типа патологий.
При снижении концентрации соматотропина человек развивается нормально, однако его рост не превышает 120 см - «гипофизарный нанизм». Такие люди (гормональные карлики) способны к деторождению и их гормональный фон не сильно нарушен.
При повышении концентрации соматотропина человек так же развивается нормально, однако его рост превышает 195 см. Такая патология называется «гигантизм»
После 20 лет выработка соматотропина снижается, следовательно, и формирование хрящевой ткани (как один из аспектов роста) замедляется и уменьшается. Поэтому костная ткань потихоньку «съедает» хрящевую ткань, следовательно кости некуда расти, кроме как в диаметре. Если выработка соматотропина не прекращается после 20, то кости начинают расти в диаметре. За счёт такого утолщения кости утолщаются например пальцы, и из-за этого утолщения они почти теряют подвижность. При этом соматотропин так же стимулирует выработку соединительной ткани, вследствие чего увеличиваются губы, нос, ушные раковины, язык и т. д. Эта патология называется «акромегалия».
Тиреотропин
Мишенью тиреотропина является щитовидная железа. Он регулирует рост щитовидной железы и выработку её основного гормона — тироксина. Пример действия релизинг-фактора: Тироксин необходим для повышения эффективности кислородного дыхания, для тироксина нужен тиреотропин, а для тиреотропина нужен тиреолиберин, который является релизинг-фактором тиреотропина.
Гонадотропины
Название гонадотропины (ГТ) обозначает два разным гормона — фолликулостимулирующий гормон и лютеинизирующий гормон. Они регулируют деятельность половых желез — гонад. Как и другие тропные гормоны, гонадотропины в первую очередь влияют на эндокринные клетки гонад, регулируя выработку половых гормонов. Кроме того, они оказывают влияние на созревание гамет, менструальный цикл и связанные с ним физиологические процессы.
Кортикотропные гормоны
Мишень КТ — кора надпочечников. Следует отметить, что паращитовидная железа регулирует минеральный обмен (с помощью парат-гормона), как и кора надпочечников, так что можно поставить регуляцию только на кору надпочечников, а паращитовидная железа автоматически будет работать в соответствии с корой надпочечников.
Эндокринная деятельность щитовидной железы. Гипоталамо-гипофизарная система регуляции эндокринной деятельности щитовидной железы.
Основной структурно-функциональной единицей щитовидной железы являются фолликулы. Фолликулы заполнены коллоидом и содержат гормоны тироксин и трийодтиронин, которые связаны с белком тиреоглобулином. В межфолликулярном пространстве находятся также С-клетки, в к-рых вырабатывается тиреокальцитонин. Действие гормонов щитовидной железы проявляется резким усилением метаболической активности организма. При этом ускоряются все виды обмена в-в , что приводит к увеличению энергообразования и повышению основного обмена.
В детском возрасте недостаток гормонов щитовидной железы приводит к задержке умственного и физического развития. У взрослых при гипофункции щитовидной железы наблюдается торможение нервно-психической активности; при избытке гормонов наблюдаются эмоциональная лабильность, возбуждение, бессонница.
Секреция гормонов щит. железы регулируется гипоталамическим тиреолиберином. Выработка тироксина и трийодтиронина резко усиливается в условиях длительного эмоционального возбуждения. Также секреция этих гормонов ускоряется при снижении температуры тела.
Йодсодержащие гормоны щитовидной железы, биосинтез и физиологическое действие йодсодержащих гормонов щитовидной железы.
Йодированные гормоны образуются в фолликулах железистой ткани, его образование происходит в три этапа:
1) образование коллоида, синтез тиреоглобулина;
2) йодирование коллоида, поступление йода в организм, всасывание в виде йодидов. Йодиды поглощаются щитовидной железой, окисляются в элементарный йод и включаются в состав тиреоглобулина, процесс стимулируется ферментом – тиреоидпероксиказой;
3) выделение в кровоток происходит после гидролиза тиреоглобулина под действием катепсина, при этом освобождаются активные гормоны – тироксин, трийодтиронин.
Роль йодированных гормонов:
1) влияние на функции ЦНС. Гипофункция ведет к резкому снижению двигательной возбудимости, ослаблению активных и оборонительных реакций;
2) влияние на высшую нервную деятельность. Включаются в процесс выработки условных рефлексов, дифференцировки процессов торможения;
3) влияние на рост и развитие. Стимулируют рост и развитие скелета, половых желез;
4) влияние на обмен веществ. Происходит воздействие на обмен белков, жиров, углеводов, минеральный обмен. Усиление энергетических процессов и увеличение окислительных процессов приводят к повышению потребления тканями глюкозы, что заметно снижает запасы жира и гликогена в печени;
5) влияние на вегетативную систему. Увеличивается число сердечных сокращений, дыхательных движений, повышается потоотделение;
6) влияние на свертывающую систему крови. Снижают способность крови к свертыванию (уменьшают образование факторов свертывания крови), повышают ее фибринолитическую активность (увеличивают синтез антикоагулянтов). Тироксин угнетает функциональные свойства тромбоцитов – адгезию и агрегацию.
Кальцитонин, паратирин, кальцитриол как компоненты системы гормональной регуляции кальциевого гомеостаза.
Кальцитонин - пептидный гормон щитовидной железы. Он получил свое название благодаря способности снижать концентрацию кальция в крови. Секретируется парафолликулярными С-клетками.
Кальцитонин - это главный антагонист паратиреодиного гормона, основным механизмом действия которого является торможение остеокластической костной резорбции.
Паратгормон
Человек имеет 2 пары околощитовидных желез, расположенных на задней поверхности или погруженных внутри щитовидной железы. Главные, или оксифильные, клетки этих желез вырабатывают паратгормон, или паратирин, или паратиреоидный гормон (ПТГ).
•паратгормон регулирует обмен кальция в организме и поддерживаетего уровень в крови.
•в костной ткани паратгормон усиливает функцию остеокластов, что приводит к деминерализации кости и повышению содержания кальция в плазме крови (гиперкальциемия).
•в почках паратгормон усиливает реабсорбцию кальция.
•в кишечнике повышение реабсорбции кальция происходит благодаря стимулирующему действию паратгормона на синтез кальцитриола –активного метаболита витамина D3.
Кальцитриол
Активная форма витамина D животных стероидной природы. Работает как сигнальная молекула. Регулирует обмен фосфата и кальция в организме.
Кальцитриол контролирует обмен кальция. В клетках кишечника он индуцирует синтез Са2±переносящих белков, которые обеспечивают всасывание ионов кальция и фосфатов из полости кишечника в эпителиальную клетку кишечника и далее транспорт из клетки в кровь против концентрационного градиента на мембранах кишечника.
В почках кальцитриол стимулирует реабсорбцию ионов кальция и фосфатов. При низкой концентрации ионов кальция кальцитриол способствует мобилизации кальция из костной ткани.
Гормоны клубочковой зоны коры надпочечников, их физиологическое действие.
Гормоны клубочковой зоны надпочечников, их функции
В клубочковой зоне происходит синтез минералокортикоидов, основным представителем которых является альдостерон.
Альдостерон усиливает в дистальных канальцах почек реабсорбцию ионов Na+, одновременно увеличивая при этом выведение с мочой ионов К+. Аналогичное усиление натрий-калиевого обмена происходит в потовых слюнных железах, а также в кишечнике. Это приводит к изменению электролитного состава плазмы крови (гипернатриемия и гипокалиемия). Кроме того, под влиянием альдостерона резко возрастает почечная реабсорбция воды, которая всасывается пассивно по осмотическому градиенту, создаваемому ионами Na+. Это приводит к существенным изменениям гемодинамики — увеличивается объем циркулирующей крови, возрастает АД. Вследствие усиленного обратного всасывания воды уменьшается диурез.
При повышенной секреции альдостерона увеличивается склонность к отекам, что обусловлено задержкой в организме натрия и воды, повышением гидростатического давления крови в капиллярах и в связи с этим — усиленной экссудацией жидкости из просвета сосудов в ткани. За счет усиления процессов экссудации и отечности тканей альдостерон способствует развитию воспалительной реакции и является провоспалительным гормоном. Под влиянием альдостерона увеличивается также секреция ионов Н+ в канальцевом аппарате почек, что приводит к снижению их концентрации во внеклеточной жидкости и изменению кислотно-основного состояния (алкалоз).
Снижение секреции альдостерона вызывает усиленное выведение натрия и воды с мочой, что приводит к дегидратации тканей, снижению объема циркулирующей крови и уровня АД. В результате в организме возникают явления циркуляторного шока. Концентрация калия в крови при этом, наоборот, увеличивается, что является причиной нарушения электрической стабильности сердца и развития сердечных аритмий.
Ренин-ангиотензин-альдостероновая система, ее физиологические функции.
Ренин-ангиотензин-альдостероновая система, ее физиологическое действие
Основным фактором, регулирующим секрецию альдостерона, является функционирование ренин-ангиотензин-альдостероновой системы. При снижении АД наблюдается возбуждение симпатической части автономной нервной системы, что приводит к сужению почечных сосудов. Уменьшение почечного кровотока способствует усиленной выработке ренина в юкстагломерулярных нефронах почек. Ренин является ферментом, который действует на плазменный α2-глобулин ангиотензиноген, превращая его в ангиотензин I. Образовавшийся ангиотензин I затем превращается в ангиотензин II, который увеличивает секрецию альдостерона. Выработка альдостерона может усиливаться также по механизму обратной связи при изменении электролитного состава плазмы крови, в частности при гипонатриемии или гиперкалиемии. В незначительной степени секреция этого гормона стимулируется кортикотропином.
Атриопептид и его роль в системе гормональной регуляции натриевого гомеостаза.
Предсердный натрийуретический пептид (ПНП) синтезируется и секретируется кардиомиоцитами предсердий, которыерые содержат гранулы гормона. Это полипептид, циркулирует в крови и имеет 28 аминокислотных остатков. Стимулирует выделение ПНП растяжение предсердий кровью при увеличении их наполнения - гиперволемии, что приводит к увеличению количества гранул в кардиомиоцитах. ПНП увеличивает выведение ионов Na+ почками при гиперволемии. Вслед за Na повышается выделение H2O с осмотическим градиентом, что ведет к нормализации объема крови и уменьшение концентрации ионов Na в крови и внеклеточной жидкости. Увеличение экскреции ионов Na почками является следствием влияния ПНП на такие функции клеток-мишеней: расширяет афферентные артериолы в клубочках нефрона, что приводит к увеличению клубочковой фильтрации и поступления ионов Na + в капсулу нефрона; подавляет реабсорбцию ионов Na + в канальцах нефрона, следствием чего является рост выведение ионов Na + с мочой и воды за осмотическим градиентом; тормозит секрецию ренина юкстагломерулярного клетками (ЮГК), в результате чего уменьшается концентрация ангиотензина II и альдостерона, что приводит к снижению реабсорбции ионов Na +.
Гормоны пучковой зоны коры надпочечников, их физиологическое действие.
Гормоны пучковой зоны коры надпочечников, их физиологические функции
Главный представитель – кортизол. Кортизол — биологически активный глюкокортикоидный гормон стероидной природы
Основные функции гормона:
Снижает распад глюкозы в мышцах и одновременно повышает ее распад в других участках организма. Это нужно, чтобы обеспечить активную работу мышц и скорость в опасных условиях (например, если придется убегать и драться).
Усиливает работу сердца и повышает сердечный ритм. При этом кровяное давление нормализуется, чтобы в момент опасности человеку не стало плохо.
Улучшает работу мозга, обостряет все мыслительные процессы, помогает сконцентрироваться на появившейся проблеме.
Подавляет любую воспалительную реакцию в организме или аллергический ответ, улучшает деятельность печени.
Особую роль играет кортизол при беременности – гормон отвечает за формирование легочной ткани у плода.
Гипоталамо-гипофизарная система регуляции эндокринной деятельности пучковой зоны коры надпочечников.
ГГС- путь характеризующийся тем, что в медиобазальном гипоталамусе вырабатываются аденогипофизотропные гормоны (либерины и статины), которые с кровью доносятся до передней доли гипофиза. Аксоны нейросекреторных клеток, вырабатывающих либерины и статины, из медиобазального гипоталамуса направляются в срединное возвышение и заканчиваются аксовазальными синапсами на капиллярах первичной сети. Через эти синапсы либерины или статины поступают в кровеносное русло этих капилляров и далее транспортируются через портальные вены во вторичную капиллярную сеть (вторичной сети впадают в выносящие вены гипофиза, т. е. эти капилляры расположены между венами (портальными и выносящими) и поэтому формируют чудесную сеть). Через стенку капилляров либерины или статины поступают в паренхиму передней доли гипофиза и захватываются рецепторами эндокринных клеток (тиролиберины захватываются тиротропными аденоцитами, гонадолиберины — гонадотропными аденоцитами и т. д.). В результате этого из аденоцитов выделяются тропные гормоны, которые поступают в капилляры вторичной сети и транспортируются с током крови к соответствующим железам.
Под влиянием кортиколиберинов (или кортикостатинов, подавляющих синтез), вырабатываемых в ядрах медиобазального гипоталамуса, эти клетки секретируют кортикотропный или адренокортикотропный гормон (АКТГ), стимулирующий функцию коры надпочечников.
Гормоны сетчатой зоны коры надпочечников, их физиологическое действие.
В сетчатом слое в норме вырабатываются половые стероиды. Основные биологически активные вещества этой зоны – дегидроэпиандростерон и андростендион. Эти вещества по своей природе являются слабыми андрогенами. Они в десятки раз слабее тестостерона.
Дегидроэпиандростерон и андростендион являются основными мужскими половыми гормонами в женском организме.
Они необходимы для:
1) формирования полового влечения;
2) поддержания либидо;
3) стимуляции работы сальных желез;
4) стимуляции роста волос в андрогензависимых зонах;
5) стимуляции появления части вторичных половых признаков;
6) формирования некоторых психологических реакций (агрессия)
7) формирование некоторых интеллектуальных функций (логика, пространственное мышление).
Тестостерон и эстрогены в надпочечниках не синтезируются. Однако из слабых андрогенов (дегидроэпиандростерона и андростендиона) на периферии (в жировой ткани) могут образовываться эстрогены.
У женщин этот путь является основным способом синтеза половых гормонов в постменопаузе. У мужчин с ожирением эта реакция может способствовать феминизации (приобретению несвойственных черт внешности и психики).
Гормоны мозгового вещества надпочечников, их физиологическое действие. Гипоталамо-симпато-адреналовая система.
Гормоны мозгового вещества надпочечников, их физиологическое действие.
В мозговом веществе надпочечников содержатся хромаффинные клетки, в которых синтезируются адреналин и норадреналин. Продукция этих гормонов резко усиливается при возбуждении симпатической части автономной нервной системы. В свою очередь выделение этих гормонов в кровь приводит к развитию эффектов, аналогичных действию стимуляции симпатических нервов. Разница состоит лишь в том, что гормональный эффект является более длительным. К наиболее важным эффектам катехоламинов относятся стимуляция деятельности сердца, вазоконстрикция, торможение перистальтики и секреции кишечника, расширение зрачка, уменьшение потоотделения, усиление процессов катаболизма и образования энергии.
Адреналин имеет большее сродство к β-адренорецепторам, локализующимся в миокарде, вследствие чего вызывает положительные инотропный и хронотропный эффекты в сердце. С другой стороны, норадреналин имеет более высокое сродство к сосудистым α-адренорецепторам. Поэтому, вызываемые катехоламинами вазоконстрикция и увеличение периферического сосудистого сопротивления в большей степени обусловлены действием норадреналина.
Адреналин выполняет функцию гормона, он поступает в кровь постоянно, при различных состояниях организма (кровопотере, стрессе, мышечной деятельности) происходит увеличение его образования и выделения в кровь.
Возбуждение симпатической нервной системы приводит к повышению поступления в кровь адреналина и норадреналина, они удлиняют эффекты нервных импульсов в симпатической нервной системе. Адреналин влияет на углеродный обмен, ускоряет расщепление гликогена в печени и мышцах, расслабляет бронхиальные мышцы, угнетает моторику ЖКТ и повышает тонус его сфинктеров, повышает возбудимость и сократимость сердечной мышцы. Он повышает тонус кровеносных сосудов, действует сосудорасширяюще на сосуды сердца, легких и головного мозга. Адреналин усиливает работоспособность скелетных мышц.
Повышение активности адреналовой системы происходит под действием различных раздражителей, которые вызывают изменение внутренней среды организма. Адреналин блокирует эти изменения.
Адреналин – гормон короткого периода действия, он быстро разрушается моноаминоксидазой. Это находится в полном соответствии с тонкой и точной центральной регуляцией секреции этого гормона для развития приспособительных и защитных реакций организма.
Норадреналин выполняет функцию медиатора, он входит в состав симпатина – медиатора симпатической нервной системы, он принимает участие в передаче возбуждения в нейронах ЦНС.
Гипоталамо-симпато-адреналовая система
Синтез катехоламинов в мозговом веществе надпочечников стимулируется нервными импульсами, поступающими из гипоталамуса к Th5-9 сегментам с.м. далее по чревному симпатическому нерву. Выделяющийся в синапсах ацетилхолин взаимодействует с холинергическими рецепторами никотинового типа и возбуждает нейросекреторную клетку надпочечника. Благодаря существованию нервно-рефлекторных связей надпочечники отвечают усилением синтеза и выделения катехоламинов в ответ на болевые и эмоциональные раздражители, гипоксию, мышечную нагрузку, охлаждение и т. д.
Механизм гипергликемического действия глюкагона. Механизм гипогликемического действия инсулина.
Интенсивность выработки гормонов поджелудочной железы контролируется вегетативной нервной системой (парасимпатические нервы вызывают гипогликемию, а симпатические - гипергликемию). Однако основными факторами регуляции секреторной активности клеток в островках Лангерганса, являются концентрации питательных веществ в крови и просвете ЖКТ. Благодаря этому, своевременные реакции клеток островкового аппарата обеспечивают поддержание постоянного уровня питательных веществ в крови между приемами корма.
Механизм гипергликемического действия глюкагона реализуется за счет его связывания со спецфическими рецепторами на поверхности клеток органов-мишеней и активации аденилатциклазы, которая катализирует превращение АТФ в цАМФ. В результате: стимуляция процессов гликогенолиза и глюконеогенеза в печени и увеличивается концентрация глюкозы в крови; Избыточное содержание инсулина в крови вызывает гипогликемию. Это может привести к потере сознания. Поглощение глюкозы мозговой тканью определяется ее концентрацией в плазме крови. Выработка инсулина регулируется механизмом отрицательной обратной связи в зависимости от концентрации глюкозы в плазме крови. Повышенное содержание глюкозы в крови способствует увеличению выработки инсулина; при гипогликемии тормозится.
Гипоталамо-гипофизарная система регуляции половых желез. Гормоны яичников, их функции. Гормоны семенников, их функции.
Гипоталамо-гипофизарная система регуляции эндокринной деятельности половых желез.
Секреция тестостерона (наиболее важный мужской половой гормон) регулируется лютеинизирующим гормоном аденогипофиза, продукция которого возрастает в период полового созревания. при увеличении содержания в крови тестостерона по механизму отрицательной обратной связи тормозится выработка лютеинизирующего гормона. Уменьшение продукции обоих гонадотропных гормонов фолликулостимулирующего и лютеинизирующего, происходит также при ускорении процессов сперматогенеза. Секреция женских половых гормонов характеризуется определенной цикличностью, связанной с изменением продукции гипофизарных гонадотропинов в течение менструального цикла. Выработка эстрогенов и прогестерона регулируется гипофизарными гонадотропинами, продукция которых возрастает у девочек, начиная с 9-10 лет. Секреция гонадотропинов тормозится при высоком содержании в крови женских половых гормонов
Гормоны яичников, их функции.
Основными гормонами яичника являются эстрогены, прогестерон и андрогены.
Эстрогены включает 3 фракции: Эстрадиол (Е2); Эстрон (Е1); Эстриол (Ез). Способствуют развитию вторичных половых признаков, регенерации и росту эндометрия в матке, стимулируют секрецию шеечной слизи, сократительную активность гладкой мускулатуры половых путей; изменяют все виды обмена в-в с преобладанием процессов катаболизма; понижают температуру тела. Гестагены способствуют нормальному развитию беременности. Основной гестагенный гормон: прогестерон. Под их влиянием подавляются возбудимость и сократительная способность миометрия при одновременном увеличении его растяжимости и пластичности; способствуют усвоению организмом в-в поступающих извне. Андрогены: В яичнике: андростендион, дегидроэпиандростерон и дегидроэпиандростерон-сульфат. Оказывают влияние на молочную железу, тормозя секрецию молока у кормящих женщин; принимают участие в регуляции белкового, жирового и водного обмена. Длительное введение андрогенов приводит к накоплению в организме P, S, Na,Сa и воды. Андрогены, вводимые в организм женщины в больших дозах, вызывают явления вирилизации: рост волос на лице и др.
Гормоны семенников, их функции
Мужские половые гормоны – андрогены (истинно мужской половой гормон - тестостерон, в меньшей степени - андростерон); наряду с андрогенами семенники вырабатывают небольшое количество женских половых гормонов – эстерогенов. Функции тестостерона: 1.Развитие вторичных половых признаков (пропорции тела, мускулатура, рост бороды, волос на теле и др.) 2.Рост и развитие органов размножения
3.Половое созревание
4.Половое поведение, половое влечение, половая потенция.
5.Участие в образовании подвижных зрелых сперматозоидов
6.Регулирует секрецию придаточных желёз (предстательной железы, семенных пузырьков)
7.Усиливает рост тела, синтез белка, уменьшают количество жира Андрогены образуются ещё до полового созревания, но не в семенниках, а в коре надпочечников.
Эндотелий кровеносных сосудов как эндокринная ткань. Физиологические эффекты биологически активных веществ, синтезируемых эндотелиальными клетками.
Эндотелий кровеносных сосудов как эндокринная ткань. Физиологические эффекты биологически активных веществ, синтезируемых эндотелиальными клетками.
Эндотелий сосудов продуцирует 2 в-ва, обладащие многими физиологическими эффектами:
1) эндотелиальный релаксирующий фактор. Он образуется из аминокислоты 1- аргинина под влиянием NO-синтазы. Также продуцируется в нейронах центральной и периферической НС и в синапсах нехолинергических, неадренергических нервных во-локон, которые вызывают расслабление миоцитов азотергических волокон. Она способна угнетать агрегацию тромбоцитов; продукцию в почках ренина, вызывать дезагрегацию агрегированных тромбоцитов, изменять а-адренореактивность миоцитов сосудов.
2) эндотелин. Основной эффект эндотелина: повышение тонуса гладких мышц сосудов. Этот пептид является одним из наиболее мощных на сегодня вазоконстрикторов. Введение эндотелина вызывает спазм сосудов и рост АД. Также повышает сердечный выброс, увеличивает тонус гладких мышц дыхательных путей, ЖКТ, матки; повышает продукцию атриопептида в миокарде, образов ренина, вазопрессина, АКТГ, кортикостерона.
Инстинкты, их роль в приспособительной деятельности человека. Классификация инстинктов, их характеристика.
Инстинкты являются врожденными, безусловными рефлексами и представляют собой совокупность двигательных актов и сложных форм поведения (пищевые, половые, самосохранения). Они имеют особенности проявления и функционирования, связанные с физиологическими свойствами:
1) морфологическим субстратом служат лимбическая система, базальные ядра, гипоталамус;
2) носят цепной характер, т. е. время окончания действия одного безусловного рефлекса является стимулом для начала действия следующего;
3) для проявления большое значение имеет гуморальный фактор (например, для пищевых рефлексов – снижение уровня глюкозы в крови);
4) имеют готовые рефлекторные дуги;
5) составляют основу для условных рефлексов;
6) передаются по наследству и носят видовой характер;
7) отличаются постоянностью и мало изменяются в течение жизни;
8) не требуют дополнительных условий для проявления, возникают на действие адекватного раздражителя.
Инстинкты, их роль в приспособительной деятельности человека.
В формировании целостного поведения животных большую роль играют инстинкты. Частично роль их сохраняется и при формировании поведения человека, но, как правило, в проявлении их доминирует мыслительная деятельность. Человек сознательно может подавлять или резко сглаживать проявления инстинктов.
Выраженность инстинкта зависит от многих факторов внешней и внутренней среды (от конкретной ситуации, метаболизма, гормональных влияний, процессов мышления). Фазы проявления инстинктивной деятельности. Строение цепочки рефлекторных актов в процессе развития инстинкта происходит в две фазы: первая-поисковая, вторая - завершающая.
В первую фазу проявления инстинктивного поведения достаточно пластичны, зависят от конкретных, реальных условиях.
Заключительная же фаза, напротив, отмечается жестким стереотипом, который зависит от анатомо-физиологических особенностей нервной системы. В отличие от первой фазы во вторую фазу приобретенные компоненты играют незначительную роль, а нередко отсутствуют совсем. Жесткая видовое единообразие инстинктов обеспечивает хранение жизненно важных функций, направленных на выживание особи и вида независимо от случайных условий среды.
Биологическое значение инстинктов не ограничивается только организацией поведения животных. Они являются эволюционной основой формирования сложных форм поведения.
Классификация инстинктов, характеристика разных видов инстинктов.
В классификации инстинктов выделяют 3 типа:
Биологические(самосохранение особи и вида): оборонительный, половой, родительский, пищевой, питьевой, территориальный, энергосберегаемый
Социальные(существование в обществе): сопереживание и сочувствие, стремление к лидерству.
Личностные(развитие личности): подражательный, игровой, самоутверждение и тд.
Условные рефлексы, их роль в приспособительной деятельности человека. Классификация условных рефлексов, их характеристика.
Условные рефлексы вырабатываются в течение жизни, так как не имеют готовых рефлекторных дуг. Они носят индивидуальный характер и в зависимости от условий существования могут постоянно меняться. Их особенности:
1) морфологическим субстратом является кора больших полушарий, при ее удалении старые рефлексы исчезают, а новые не вырабатываются;
2) на их базе формируется взаимодействие организма с внешней средой, т. е. они уточняют, усложняют и делают тонкими данные отношения.
Биологическое значение условных рефлексов заключается в резком расширении числа сигнальных, значимых для организма раздражителей, что обеспечивает несравненно более высокий уровень адаптивного (приспособительного) поведения.
Условно-рефлекторный механизм лежит в основе формирования любого приобретенного навыка, в основе процесса обучения. Структурно-функциональной базой условного рефлекса служат кора и подкорковые образования мозга.
Сущность условно-рефлекторной деятельности организма сводится к превращению индифферентного раздражителя в сигнальный, значащий, благодаря многократному подкреплению раздражения безусловным стимулом. Благодаря подкреплению условного стимула безусловным ранее индифферентный раздражитель ассоциируется в жизни организма с биологически важным событием и тем самым сигнализирует о наступлении этого события. При этом в качестве эффекторного звена рефлекторной дуги условного рефлекса может выступать любой иннервируемый орган. В организме человека и животных нет органа, работа которого не могла бы измениться под влиянием условного рефлекса. Любая функция организма в целом или отдельных его физиологических систем может быть модифицирована (усилена или подавлена) в результате формирования соответствующего условного рефлекса.
Классификация условных рефлексов, характеристика разных видов условных рефлексов.
По отношению условного сигнала к вызываемой им реакции различают натуральные и искусственные условные рефлексы.
Натуральными называют условные рефлексы, которые образуются на раздражители, являющиеся естественными признаками безусловного стимула, на базе которого они вырабатываются. Натуральные условные рефлексы по сравнению с искусственнымиотгличаются большей легкостью образования и большей прочностью
Искусственными называют условные рефлексы образующиеся на стимулы, которые обычно не имеют прямого отношения подкрепляющему их безусловному стимулу,
В зависимости от природы рецепторных структур, на которые действу ют условные стимулы, различают экстероцептивные, интероцептивные и проприоцептивные условные рефлексы
Экстероцетпивные условные рефлексы образуемые на стимулы, воспринимаемые наружными рецепторами тела, составляют основную массу условнорефлекторных реакций, обеспечивающих адаптивное поведение животных и человека в условиях изменяющееся внешней среды.
Интероцептивные условные рефлексы вырабатываемые, на раздражения интерорецепторов, обеспечивают физиологические процессы гомеостатической регуляции функции внутренних органов.
Проприоцептивные условные рефлексы, формируемые на раздражение собственных рецепторов скелетных мышц туловища и конечностей, со ставляютосновувсex двигательных навыков животных и человека.
В зависимости от структуры применяемого условного стимула различают простые и сложные (комплексные) условные рефлексы
В случае простого условного рефлекса в качестве условного стимула используется простой раздражитель.
В реальных условиях функционирования организма в качестве условных сигналов выступают, как правило, не отдельные раздражители, а их временные и пространственные комплексы. В это случае в качестве условного стимула выступает либо вся окружающая животное обстановка, либо части ее в виде комплекса сигналов
Одной из разновидностей такого комплексного условного рефлекса является стереотипный условный рефлекс, образуемый на определенный временной или пространственный комплекс стимулов.
Различают также условные рефлексы, вырабатываемые на одновременные и последовательные комплексы стимулов, на последовательную цепь условных раздражителей, разделенных определенным временным промежутком
Следовые условные рефлексы формируются в том случае, когда безусловный подкрепляющий раздражитель предъявляется лишь после окончания действия условного стимула
Различают условые рефлексы первого, второго, третьего и т.д. порядкa.
Если условный стимул подкрепляется безусловным, образуется условный рефлекс первого порядка
Условный рефлекс второго порядка образуется, если условный стимул (например, свет) подкрепляется не безусловным, а условным раздражителем, на который ранее был образован условный рефлекс.
Условные рефлексы третьего и более сложного порядка образуются труднее и отличаются меньшей прочностью. К условным рефлексам второго и более высокого порядка относятся вырабатываемые на словесный сигнал (слово представляет здесь сигнал, на который ранее был образован условный рефлекс при подкреплении его безусловным стимулом).
Нейрофизиологический механизм образования условного рефлекса.
В зоне коркового представительства условного стимула и коркового (или подкоркового) представительства безусловного стимула формируются два очага возбуждения. Очаг возбуждения, вызванный безусловным стимулом внешней или внутренней среды организма, как более сильный (доминантный) притягивает к себе возбуждение из очага более слабого возбуждения, вызванного условным стимулом. После нескольких повторных предъявлений условного и безусловного раздражителей между этими двумя зонами появляется устойчивый путь движения возбуждения: от очага, вызванного условным стимулом, к очагу, вызванному безусловным стимулом. В результате изолированное предъявление только условного стимула теперь приводит к реакции, вызываемой ранее безусловным стимулом. В качестве главных клеточных элементов центрального механизма образования условного рефлекса выступают вставочные и ассоциативные нейроны коры большого мозга.
Правила образования, стадии образования и общие свойства условных рефлексов.
1. Для образования условного рефлекса необходимо совпадение по времени, т. е. сочетание какого-либо индифферентного условного раздражителя с раздражителем, вызывающим безусловный рефлекс.
2. Для более быстрого образования временных связей необходимо, чтобы действие условного раздражителя несколько предшествовало действию безусловного.
3. Условный раздражитель должен быть физиологически более слабым, по сравнению с безусловным, и возможно более индифферентным, т. е. не вызывающим значительной самостоятельно протекающей реакции.
4. Скорость образования условных рефлексов очень сильно зависит от степени значимости безусловного раздражителя для данного животного, т. е. безусловный раздражитель должен быть значимым.
5. Для образования условного рефлекса необходимо нормальное, деятельное состояние головного мозга.
6. Во время образования условного рефлекса должны отсутствовать посторонние раздражители, т. е. такие, которые вызывают собственные ответные реакции. Наиболее часто встречается ориентировочная реакция, при которой прекращается вся текущая деятельность собаки.
Свойства:
1. Их приспособительный характер. Смысл условного рефлекса в том, что он делает поведение особенно пластичным, подогнанным к конкретным условиям среды.
2. Любые условные рефлексы образуются при участии высших отделов головного мозга. Поэтому приспособительные реакции простейших или кишечнополостных не могут быть отнесены к реакциям условно-рефлекторного типа.
3. Условные рефлексы приобретаются и отменяются в индивидуальной жизни каждой конкретной особи. Этим они принципиально отличаются от безусловных рефлексов, одинаково проявляющихся у всех особей данного вида и в определенной мере детерминированных генотипически. Основная масса условных рефлексов образуется только при повторяемости сочетаний условного и безусловного раздражителей.
4. Условный рефлекс имеет сигнальный характер, то есть всегда предшествует, предупреждает последующее возникновение безусловного рефлекса. Смысл условного рефлекса в том, чтобы обеспечить подготовку организма к какой-либо биологически целенаправленной деятельности.
Стадии образования условных рефлексов.
Три основные стадии:
Стадия прегенерализации - кратковременная фаза, которая характеризуется выраженной концентрацией возбуждения и отсутствием условных поведенческих реакций.
Стадия генерализации. Это феномен, который возникает на начальных этапах выработки условного рефлекса. Требуемая реакция в этом случае вызывается не только подкрепляемым стимулом, но и другими, более или менее близкими к нему. Во время стадии генерализации условные реакции возникают на сигнальные и другие раздражители (явление афферентной генерализации), а также в интервалах между предъявлениями условного стимула. В основе этой стадии лежит иррадиация возбуждения. В этот период по коре и подкорковым структурам возникают биоэлектрические сдвиги (блокада альфа-ритма, десинхронизация, возникновение тета-ритма и вызванных потенциалов). Кроме этого наблюдается синхронизация биоэлектрической активности во многих участках коры и подкорковых структур - дистантная синхронизация биопотенциалов.
3. Стадия специализации. В этот период реакция возникает только на сигнальный раздражитель и объем распространения биопотенциалов уменьшается. Первоначально И.П.Павлов предполагал, что условный рефлекс образуется на уровне "кора -подкорковые образования". В более поздних работах он объяснял образование условно-рефлекторной связи образованием временной связи между корковым центром безусловного рефлекса и корковым центром анализатора. В качестве главных клеточных элементов механизма образования условного рефлекса в этом случае выступают вставочные и ассоциативные нейроны коры больших полушарий, а в основе замыкания временной связи лежит процесс доминантного взаимодействия между возбужденными центрами. Данные современной нейрофизиологии указывают на возможность разных уровней замыкания: "кора-кора", "кора-подкорковые образования", "подкорковые образования-подкорковые образования". Э.А.Асратян, изучая безусловный рефлекс, высказал гипотезу о структуре условного рефлекса как процесса синтеза безусловный рефлексов.
Торможение условных рефлексов, его роль в приспособительной деятельности человека. Классификация торможения условных рефлексов.
В зависимости от природы физиологического механизма, лежащего в основе тормозного эффекта на условнорефлекторную деятельность организма, различают безусловное (внешнее и запредельное) и условное (внутреннее) торможение условных рефлексов.
Внешнее торможение условного рефлекса возникает под действием другого постороннего условного или безусловного раздражителя. При этом основная причина подавления условного рефлекса не зависит от самого тормозимого рефлекса и не требует специальной выработки. Внешнее торможение наступает при первом предъявлении соответствующего сигнала.
Запредельное торможение условного рефлекса развивается либо при чрезмерно большой силе стимула, либо при низком функциональном состоянии ЦНС, на уровне которого обычные пороговые раздражители приобретают характер чрезмерных.
Условное (внутреннее) торможение условного рефлекса требует специальной выработки. Поскольку развитие тормозного эффекта связано с нейрофизиологическим механизмом образования условного рефлекса, такое торможение относится к категории внутреннего, а проявления его связаны с определенными условиями.
Биологический смысл внутреннего торможения условных рефлексов состоит в том, что изменившиеся условия внешней среды требует соответствующего приспособительного изменения в условнорефлекторном поведении. Условный рефлекс угнетается, поскольку перестает быть сигналом, предвещающим появление безусловного стимула.
Различают 4 вида внутреннего торможения: угасание, дифференцировка, условный тормоз, запаздывание.
Если условный раздражитель предъявляется без подкрепления безусловным, то через некоторое время после изолированного применения условного стимула реакция на него угасает. Такое торможение условного рефлекса называется угасательным.
Если у животного или человека с выработанным условным рефлексом на определенную частоту звукового стимула (например, звук метронома с частотой 50 в секунду) близкие по смыслу раздражители (звук метронома с частотой 45 или 55 в секунду) не подкреплять безусловным стимулом, то условнорефлекторная реакция на последние подавляется. Такой вид внутреннего торможения называют дифференцировочным. Оно лежит в основе многих форм обучения, связанных с выработкой тонких навыков.
Если условный стимул, на который образован условный рефлекс, применяется в комбинации с некоторым другим стимулом и их комбинация не подкрепляется безусловным стимулом, наступает торможение условного рефлекса, вызываемого этим стимулом. Этот вид условного торможения называется условным тормозом.
Запаздывательное торможение наступает тогда, когда подкрепление условного сигнала безусловным раздражителем осуществляется с большим опозданием(2—3мин) по отношению к моменту предъявления условного раздражителя
Типы высшей нервной деятельности по И.П. Павлову, их соотношение с типами темперамента по Гиппократу.
Систематизация типов ВНД основана на оценке трех основных особенностей процессов возбуждения и торможения: силы, уравновешенности и подвижности, выступающих как результат унаследованных и приобретенных индивидуальных качеств нервной системы. Тип как совокупность врожденных и приобретенных свойств нервной системы, определяющих характер взаимодействия организма и среды, проявляется в особенностях функционирования физиологических систем организма и прежде всего самой нервной системы, ее высших «этажей», обеспечивающих ВНД.
Типы ВНД формируются на основе как генотипа, так и фенотипа. Генотип формируется в процессе эволюции под влиянием естественного отбора, обеспечивая развитие наиболее приспособленных к окружающей среде индивидов. Под влиянием реально действующих на протяжении индивидуальной жизни условий внешней среды генотип формирует фенотип организма.
Современные представления о типах ВНД в значительной степени могут отождествляться с четырьмя типами человеческого темперамента (холерический, меланхолический, флегматический, сангвинический), выдленными еще древнегреческим врачом Гиппократом (IV в. до нашей эры) на основе наблюдения за поведением людей. Сложная комбинация передаваемых по наследству особенностей в сочетании с большим разнообразием индивидуально приобретенного поведения позволяет лишь в самых общих чертах идентифицировать определенный тип ВНД.
В условнорефлекторной деятельности сила процесса возбуждения определяется скоростью и прочностью выработки условных рефлексов, сила процесса торможения находит отражение в скорости и прочности выработки дифференцировочного и запаздывающего торможения. Подвижность нервных процессов оценивается в показателях прочности переделки сигнального значения условных раздражителей (с возбудительного на тормозной и наоборот).
Сангвинический тип характеризуется достаточной силой и подвижностью возбудительного и тормозного процессов (сильный, уравновешенный, подвижный).
Флегматический тип отличается достаточной силой обоих нервных процессов при относительно низких показателях их подвижности (сильный, уравновешенный, инертный).
Холерический тип характеризуется высокой силой возбудительного процесса с явным преобладанием его над тормозным и повышенной подвиж-
Меланхолический тип характеризуется явным преобладанием тормозного процесса над возбудительным и их низкой подвижностью (слабый, неуравновешенный, инертный)
Психонервная память, ее роль в жизнедеятельности человека. Теории механизма краткосрочной и долгосрочной памяти.
Психонервная память, ее роль в жизнедеятельности человека.
Определяющее значение памяти: она обеспечивает единство и целостность сознания и личности. Базовая функция памяти - «нанизывать» образования на один стержень. Основной объект для памяти - прошлое ==> основная функция памяти - фиксировать воспроизводить, работать с прошлым, изменяя тем самым поведение личности.
Иногда память может подменять восприятие и мышление; это проявляется в установках, стереотипах, иллюзиях и других ошибках памяти.
Память лежит в основе способностей человека, является условием научения, приобретения знаний, формирования учений и навыков. Без памяти невозможно нормальное функционирование ни личности, ни общества. Благодаря своей памяти, ее совершенствованию человек выделился из животного царства и достиг тех высот, на которых сейчас находится.
Виды психонервной памяти, их характеристика.