

Виды хеморецепторов
1. Экстероцептивные. Воспринимают информацию определенных сигналов организма.
Данный вид делится на: • вкусовые; • обонятельные.
Первые расположены на луковицах языка и реагируют на вещества, растворенные в жидкости. Вторые находятся на эпителии носовой полости, вступают во взаимодействие с веществами, растворенными в газах.
По строению: • Первичные, элементом является терминали афферентных нервных волокон. • Вторичные, элементом является специальная структура, которая не относится к нервной ткани.
2. Интероцептивные. Воспринимают сигналы жидких сред организма. Они концентрируются на рефлексогенных участках и диффузно распределяются по органам и тканям.
Общий план строения синапсов. Классификация синапсов.
Синапсы состоят из трех основных элементов: пресинаптической мембраны, постсинаптической мембраны и синаптической щели (рис. 1).

Рис. 1. Строение синапса: 1 — микротрубочки; 2 — митохондрии; 3 — синаптические пузырьки с медиатором; 4 — пресинаптическая мембрана; 5 — постсинаптическая мембрана; 6 — рецепторы; 7 -синаптическая щель
Некоторые элементы синапсов могут иметь и другие названия. Например, синаптическая бляшка — это синапс между нейронами, концевая пластинка — постсинаптическая мембрана нервно-мышечного синапса, моторная бляшка — пресинаптическое окончание аксона на мышечном волокне.

Пресинаптическая мембрана покрывает расширенное нервное окончание, которое представляет собой нейросекреторный аппарат. В пресинаптической части находятся пузырьки и митохондрии, обеспечивающие синтез медиатора. Медиаторы депонируются в гранулах (пузырьках).
Постсинаптическая мембрана - утолщенная часть мембраны клетки, с которой контактирует пресинаптическая мембрана. Она имеет ионные каналы и способна к генерации потенциала действия. Кроме того, на ней расположены специальные белковые структуры — рецепторы, воспринимающие действие медиаторов.
Синаптическая щель представляет собой пространство между пресинаптической и постсинаптической мембранами, заполненное жидкостью, близкой по составу к плазме крови.

27.Классификация синапсов
1. По местоположению и принадлежности соответствующим структурам:
периферические(нервно-мышечные, нейросекреторные, рецепторнонейрональные);
центральные(аксо-соматические, аксо-дендритные, аксо-аксональные, сомато-дендритные. сомато-соматические);
2. По эффекту действия:
возбуждающие
тормозные
3. По способу передачи сигналов:
Электрические,
химические,
смешанные.
4. По медиатору:
холинергические,
адренергические,
серотонинергические,
глицинергически. и т.д.
Механизм проведения возбуждения в электрических и химических синапсах нервной системы. Постсинаптические потенциалы в нервных синапсах, их природа.
Проведение возбуждения в электрических синапсах
Электрический ток, возникающий между пресинаптической и постсинаптической мембраной, раздражает постсинаптическую мембрану и вызывает генерацию в ней ПД. Процесс начинается с открывания Nа+-канала. Ионы Na+устремляются в клетку (по градиенту концентрации), что вызывает локальное обращение знака мембранного потенциала. При этом Na+-каналы тотчас закрываются, т. е. поток ионов Na+ в клетку длится очень короткое время. В связи с изменением мембранного потенциала открываются (на несколько мс) потенциал-управляемые К+-каналы и ионы К+ устремляются в обратном направлении, из клетки. В результате развития ПД происходит реверсия заряда пресинаптической мембраны.
Проведение возбуждения в химических синапсах
Передача возбуждения в химическом синапсе – сложный физиологический процесс, протекающий в несколько этапов. На пресинаптической мембране осуществляется трансформация электрического сигнала в химический, который на постсинаптической мембране снова трансформируется в электрический сигнал.
Синтез медиатора
Медиатором (посредником) называют химическое вещество, которое обеспечивает одностороннюю передачу возбуждения в химическом синапсе. Некоторые медиаторы (например, ацетилхолин) синтезируются в цитоплазме синаптического окончания, и там же молекулы медиатора депонируются в синаптических пузырьках. Ферменты, необходимые для синтеза медиатора, образуются в теле нейрона и доставляются в синаптическое окончание путем медленного (1–3 мм/сут) аксонного транспорта. Другие медиаторы (пептиды и др.) синтезируются и упаковываются в везикулы в теле нейрона, готовые синаптические пузырьки доставляются в синаптичекую бляшку за счет быстрого (400 мм/сут) аксонного транспорта. Синтез медиатора и образование синаптических пузырьков осуществляется непрерывно.
Секреция медиатора
Содержимое синаптических пузырьков может выбрасываться в синаптическую щель путем экзоцитоза. При опорожнении одного синаптического пузырька в синаптичекую щель выбрасывается порция (квант) медиатора, которая включает около 10000 молекул.
Для активации экзоцитоза необходимы ионы Са++ . В состоянии покоя уровень Са++ в синаптическом окончании низок и выделения медиатора практически не происходит. Приход в синаптическое окончание возбуждения приводит к деполяризации пресинаптической мембраны и открытию потенциалчувствительных Са++ -каналов. Ионы Са++ поступают в цитоплазму синаптического окончания (рис. 3, А,Б) и активируют опорожнение синаптических пузырьков в синаптическую щель (рис. 3, В).
|
Рис. 3. Передача сигнала в возбуждающем химическом синапсе. А - Д – последовательность процессов при срабатывании химического синапса; Е – деполяризация постсинаптической мембраны (ВПСП). 1 – пресинаптическая мембрана, 2 – синаптическая щель, 3 – постсинаптическая мембрана |


Взаимодействие медиатора с рецепторами постсинаптической мембраны
Молекулы медиатора диффундируют через синаптическую щель и достигают постсинаптической мембраны, где связываются с рецепторами хемочувствительных Na+ -каналов (рис. 3,Г). Присоединение медиатора к рецептору приводит к открытию Na+ -каналов, через которые в клетку входят ионы Na+ (рис. 3,Д). В результате входа в клетку положительно заряженных ионов происходит локальная деполяризация постсинаптической мембраны, которую называют возбуждающий постсинаптический потенциал (ВПСП) (рис. 3,Е).
Инактивация медиатора
Ферменты, находящиеся в синаптической щели, разрушают молекулы медиатора. В результате происходит закрытие Na+ -каналов и восстановление МП постсинаптической клетки. Некоторые медиаторы (например, адреналин) не разрушаются ферментами, а удаляются из синаптической щели путем быстрого обратного всасывания (пиноцитоза) в синаптическое окончание.
Генерация ПД
В нейро-нейрональном синапсе амплитуда ВПСП значительно меньше и недостаточна для того, чтобы деполяризовать мембрану нейрона. Поэтому для генерации ПД в нервной клетке требуется возникновение нескольких ВПСП. ВПСП, образовавшиеся в результате срабатывания разных синапсов, электротонически распространяются по мембране клетки, суммируются и генерируют образование ПД в области аксонного холмика. Мембрана нейрона в области аксонного холмика обладает низким электрическим сопротивлением и имеет большое количество потенциалчувствительных Na+ -каналов
Постсинаптические потенциалы в нервных синапсах, их природа
В возбуждающих синапсах нервной системы медиатором может являться ацетилхолин, норадреналин, дофамин, серотонин, глугаминовая кислота, вещество Р, а также большая группа других веществ, являющихся, если не медиаторами в прямом значении, то во всяком случае модуляторами (меняющими эффектиьность) синаптической передачи. Возбуждающие медиаторы вызывают появление на постсинаптической мембране возбуждающего постсинаптического потенциала (ВПСП). Его формирование обусловлено тем, что медиатор-рецепторный комплекс активирует Na- каналы мембраны (а также вероятно и Са-каналы) и вызывает за счет поступления натрия внутрь клетки деполяризацию мембраны. Одновременно происходит и уменьшение выхода из клетки ионов К+Амплитуда одиночного ВПСП однако довольно мала, и для уменьшения заряда мембраны до критического уровня деполяризации необходима одновременная активация нескольких возбуждающих синапсов.
ВПСП, образуемые на постсинаптической мембране этих синапсов, способны суммироваться, т.е. усиливать друг друга, приводя к росту амплитуды ВПСП (пространственная суммация).
Растет амплитуда ВПСП и при увеличении частоты поступающих к синапсу нервных импульсов (временная суммация), что повышает число выводимых в синаптическую щель квантов медиатора.
Процесс спонтанной регенеративной деполяризации возникает в нейроне обычно в месте отхождения от тела клетки аксона, в так называемом аксонном холмике, где аксон еше не покрыт миелином и порог возбуждения наиболее низкий. Таким образом, ВПСП, возникающие в разных участках мембраны нейрона и на его дендритах, распространяются к аксонному холмику, где суммируются, деполяризуя мембрану до критического уровня и приводя к появлению потенциала действия.
Тормозной постсинаптический потенциал (ТПСП)В тормозных синапсах обычно действуют другие, тормозные, медиаторы. Среди них хорошо изученными являются аминокислота глицин (тормозные синапсы спинного мозга), гамма-аминомасляная кислота (ГАМК) — тормозной медиатор в нейронах головного мозга. Вместе с тем, тормозной синапс может иметь тот же медиатор, что и возбуждающий, но иную природу рецепторов постсинаптической мембраны. Так, для ацетилхолина, биогенных аминов и аминокислот на постсинаптической мембране разных синапсов могут существовать как минимум два типа рецепторов, и, следовательно, разные медиатор-рецепторные комплексы способны вызывать различную реакцию хемочувствительных рецепторуправляемых каналов. Для тормозного эффекта такой реакцией может являться активация калиевых каналов, что вызывает увеличение выхода ионов калия наружу и гиперполяризацию мембраны. Аналогичный эффект во многих тормозных синапсах имеет активация каналов для хлора, увеличивающая его транспорт внутрь клетки. Возникающий при гиперполяризации сдвиг мембранного потенциала получил название тормозного постсинаптического потенциала (ТПСП). На рис.3.5 показаны отличительные черты ВПСП и ТПСП. Увеличение частоты нервных импульсов, приходящих к тормозному синапсу, также как и в возбуждающих синапсах, вызывает нарастание числа квантов тормозного медиатора, выделяющихся в синаптическую щель, что, соответственно, повышает амплитуду гиперполяризационного ТПСП. Вместе с тем, ТПСП не способен распространяться по мембране и существует только локально.
В результате ТПСП уровень мембранного потенциала удаляется от критического уровня деполяризации и возбуждение становится либо вообще невозможным, либо для возбуждения требуется суммация значительно больших по амплитуде ВПСП, т.е. наличие значительно больших возбуждающих токов. При одновременной активации возбуждаюших и тормозных синапсов резко падает амплитуда ВПСП, так как деполяризующий поток ионов Na+ компенсируется одновременным выходом ионов К+ в одних видах тормозных синапсов или входом ионов СГ в других, что называют шунтированием ВПСП.
|
МПП — мембранный потенциал покоя. Стрелки у синапсов показывают направление тока. |
 Рис.3.5.
Возбуждающий (В) и тормозный (Т) синапсы
и их потенциалы.
Рис.3.5.
Возбуждающий (В) и тормозный (Т) синапсы
и их потенциалы.
Под влиянием некоторых ядов может происходить блокада тормозных синапсов в нервной системе, что вызывает безудержное возбуждение многочисленных рефлекторных аппаратов и проявляется в виде судорог. Так действует стрихнин, конкурентно связывающий рецепторы постсинаптической мембраны и не позволяющий им взаимодействовать с тормозным медиатором. Столбнячный токсин, нарушающий процесс освобождения тормозного медиатора, также угнетает тормозные синапсы.
Механизм проведения возбуждения в нервно-мышечных синапсах. Роль Ca2+ в механизме синаптического процесса.
Проведение возбуждения в нервно-мышечных синапсах
ПД достигая нервного окончания (пресинаптической мембраны) вызывает его деполяризацию. При деполяризации пресинаптической мембраны кальций входит в пресинаптическую терминаль через специфические потенциалозависимые кальциевые каналы в этой мембране. Увеличение концентрации кальция в нервном окончании способствует освобождению ацетилхолина, который выходит в синаптическую щель. Медиатор достигает постсинаптической мембраны и связывается там с рецепторами. В результате внутрь постсинаптической мембраны поступают ионы натрия и эта мембрана частично деполяризуется, т.е. возбуждение пока еще не распространяется дальше, а находится в синапсе. Частичная деполяризация постсинаптической мембраны называется возбуждающим постсинаптическим потенциалом (ВПСП).
Под влиянием ВПСП в соседнем чувствительном участке мембраны мышечного волокна возникает распространяющийся ПД, который и вызывает сокращение мышцы.
Для восстановления возбудимости постсинаптической мембраны после очередного импульса необходима инактивация медиатора (инактивационная система). В противном случае, при длительном действии медиатора происходит снижение чувствительности рецепторов к этому медиатору.
Для заблокирования передачи возбуждения через синапс применяют яд кураре, который связывается с рецепторами постсинаптической мембраны и препятствует их взаимодействию с ацетилхолином. Заблокировать проведение возбуждения через синапс может яд бутулин и другие вещества.
Роль кальция в синаптических процессах
Слияние синаптических пузырьков с пресинаптической мембраной происходит, когда увеличивается концентрации Са2+ в цитозоле нервной терминали. Белок синаптического пузырька синаптотагмин связывается с Са2+ и тем самым принимает участие в регуляции экзоцитоза (в том числе реорганизуя примембранный цитоскелет).

Торможение в нервной системе. Виды торможения. Природа пре-, постсинаптического, возвратного и пессимального торможения.
Торможение - местный нервный процесс, приводящий к угнетению или предупреждению
возбуждения. Торможение является активным нервным процессом, результатом которого
служит ограничение или задержка возбуждения. Одна из характерных черт тормозного
процесса- отсутствие способности к активному распространению по нервным структурам.
Постсинаптическое торможение —основной вид торможения, заключается в том, что в нервных окончаниях тормозящих нейронов под влиянием приходящего по аксону импульса выделяется медиатор, который гиперполяризует постсинаптическую мембрану другого возбуждающего нейрона. В мембране последнего деполяризация не может достигнуть критического уровня, и поэтому возбуждение по нейрону не распространяется.
Пресинаптическое торможение локализуется в пресинаптических окончаниях, т.е. в разветвлениях (терминалях) возбуждающего нейрона. На этих терминалях располагаются окончания аксона тормозящего нейрона. При его возбуждении тормозной медиатор частично или полностью блокирует проведение возбуждения возбуждающего нейрона, и его влияние не передается на другой нейрон.
Возвратное торможение осуществляется вставочными тормозными клетками (клетками Реншоу). Аксоны мотонейронов часто дают коллатерали (ответвления), оканчивающиеся на клетках Реншоу. Аксоны клеток Реншоу оканчиваются на теле или дендритах этого мотонейрона, образуя тормозные синапсы. Возбуждение, возникающее в мотонейроне, распространяется по прямому пути к скелетной мускулатуре, а также по коллатералям к тормозящему нейрону, который посылает импульсы к мотонейронам и тормозит их. Чем сильнее возбуждение мотонейрона, тем сильнее возбуждаются клетки Реншоу и тем более интенсивно они оказывают свое тормозящее действие, что предохраняет нервные клетки от перевозбуждения.
Рис.9.
Механизмы и виды торможения 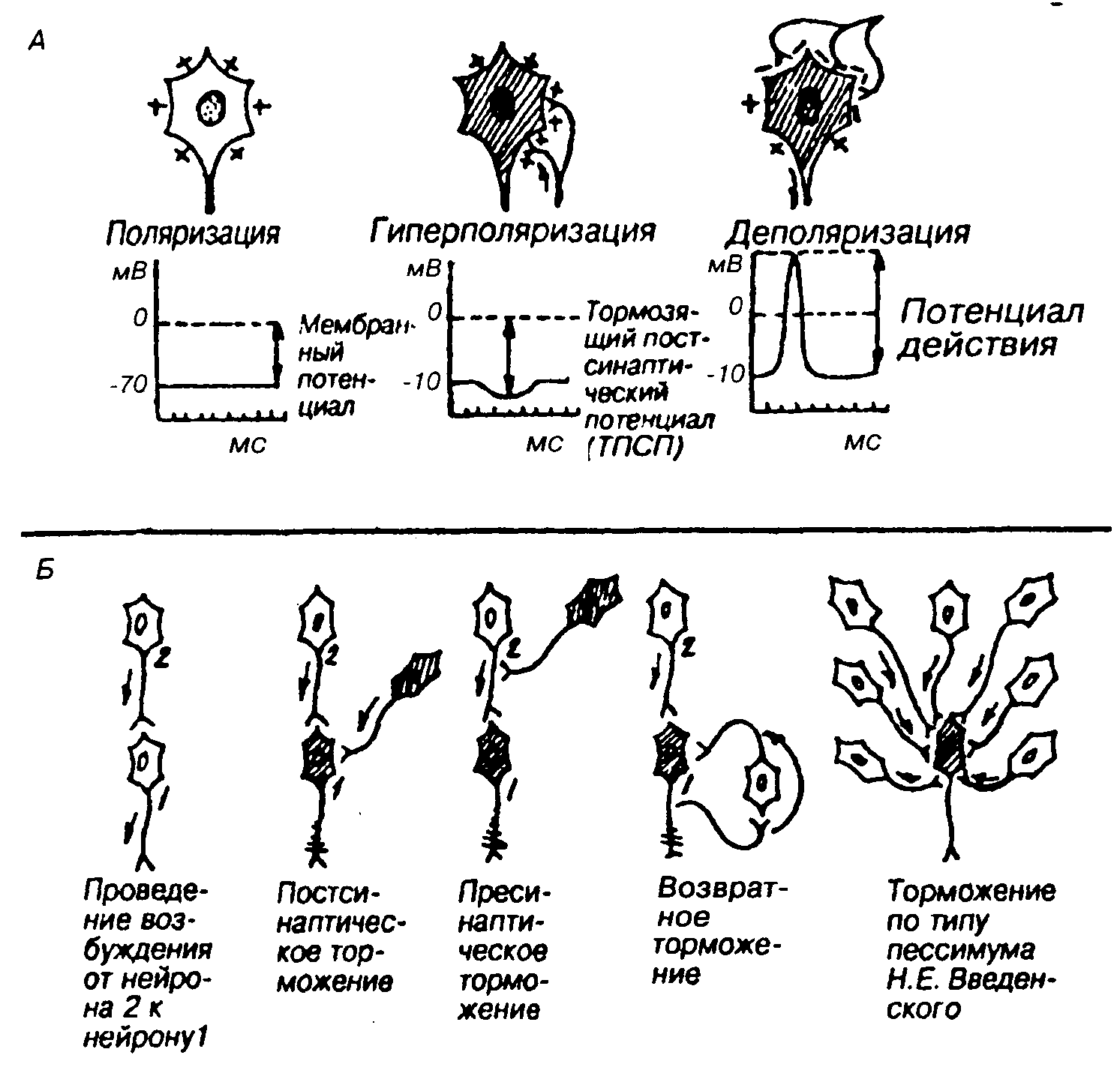 Возможные
состояния нейрона(А) и виды
внутрицентрального торможения (Б)
Возможные
состояния нейрона(А) и виды
внутрицентрального торможения (Б)
Пессимальное торможение — торможение, которое развивается в возбуждающих синапсах в результате сильной деполяризации постсинаптической мембраны под действием множественной импульсации.
Классификация рецепторов. Общие механизмы возбуждения рецепторов, биоэлектрические явления в них (рецепторный и генераторный потенциалы).
По характеру ощущений, возникающих при их раздражении: зрительные, слуховые, обонятельные, вкусовые, осязательные рецепторы, термо-, проприо- и вестибулорецепторы и рецепторы боли. По расположению: экстерорецепторы и интерорецепторы. К экстерорецепторам относятся слуховые, зрительные, обонятельные, вкусовые, осязательные. К интерорецепторам относятся вестибуло- и проприорецепторы, а также висцерорецепторы. По характеру контакта со средой: дистантные, получающие информацию на расстоянии от источника раздражения (зрительные, слуховые и обонятельные), и контактные - возбуждающиеся при непосредственном соприкосновении с раздражителем (вкусовые, тактильные). По природе раздражителя: 1- фоторецепторы, механорецепторы; 2 - хеморецепторы; 3 - терморецепторы; 4 - болевые (ноцицептивные) рецепторы. По механизму активации: первично-чувствующие (обоняния, тактильные и проприорецепторы) и вторично- чувствующие (вкуса, зрения, слуха, вестибулярного аппарата). Механизм возбуждения рецепторов. 1) взаимодействие стимула с рецепторной белковой молекулой, которая находится в составе клеточной мембраны рецепторной клетки; 2) внутриклеточные процессы усиления и передачи сенсорного стимула в пределах рецепторной клетки; 3) открывание находящихся в мембране рецептора ионных каналов, через которые начинает течь ионный ток: Na+ - деполяризация, К+-гиперполяризация клеточной мембраны рецепторной клетки (возникновение рецепторного потенциала). В первично-чувствующих рецепторах этот потенциал действует на наиболее чувствительные участки мембраны, способные генерировать ПД — электрические нервные импульсы. Во вторично-чувствующих рецепторах рецепторный потенциал вызывает выделение квантов медиатора из пресинаптического окончания рецепторной клетки. Медиатор (например, АХ), воздействуя на постсинаптическую мембрану первого нейрона, изменяет ее поляризацию (генерируется постсинаптический потенциал). Постсинаптический потенциал первого нейрона сенсорной системы называют генераторным потенциалом, так как он вызывает генерацию импульсного ответа. В первично- чувствующих рецепторах рецепторный и генераторный потенциалы — одно и то же.
Биоэлектрические явления в рецепторах (рецепторный и генераторный потенциалы). Рецепторный потенциал — изменение напряжения, возникающее в рецепторе при действии адекватного стимула вследствие изменения ионной проницаемости рецепторной мембраны, градуально зависящее от интенсивности стимула.
Генераторный потенциал — изменение напряжения в рецепторе, возникающее вследствие распространения рецепторного потенциала к центральным и проксимальным отделам рецепторной клетки и порождающее потенциалы действия (нервные импульсы).
В первичночувствующих рецепторах рецепторный потенциал возникает в дистальных отделах (терминальных ветвлениях) дендрита, генераторный потенциал — в области аксонного холмика (зрительные, обонятельные рецепторы) или в первом перехвате Ранвье афферентного волокна (соматические рецепторы).
Во вторичночувствующих рецепторах рецепторный потенциал возникает в рецепторной клетке, а генераторный потенциал — в окончании афферентного волокна. Некоторые авторы в этом случае считают, что генераторный потенциал является совокупностью процессов, протекающих в базальной части рецепторной клетки и приводящих к выбросу медиатора, воздействующего на мембрану афферентного волокна.
Несмотря на то, что рецепторный и генераторный потенциал в ряде случаев совпадают по принадлежности (первичные рецепторы), они различаются по локализации и природе возникновения: причиной появления рецепторного потенциала является непосредственно сенсорное воздействие, генераторный потенциал возникает вследствие распространения рецепторного потенциала, не зависит от других факторов и развивается в области, где возможна генерация распространяющихся потенциалов .
Свойства рецепторных потенциалов:
1. Образуются в месте действия стимула.
2. Градуальность: стимулами разной интенсивности большинство рецепторов деполяризуется (или гиперполяризуется, как в случае палочек и колбочек) неодинаково. Хотя амплитуда потенциала определенным образом отражает силу стимуляции, последняя не служит источником энергии для такого изменения клетки. Единственная функция стимула - управление ионными токами через мембрану.
3. Локальность: распространяется по клетке электротонически, а не проводится активно по мембране.
4. Рецепторные потенциалы могут подвергаться пространственной и временной суммации. Таким образом, у рецепторных потенциалов много общих свойств с локальными синаптическими токами центральных нейронов.
Свойства генераторных потенциалов:
1. Образуются в месте генерации потенциала действия (аксонный холмик, первый перехват Ранвье).
2. Является результатом пространственной и временной суммации рецепторных потенциалов.
3. Градуален: рецепторным потенциалам (стимулам) разной интенсивности соответствует большее или меньшее значение. Большей величине генераторного потенциала соответствует большая частота формирования потенциала действия (следования импульсов).
4. Локальный, но является источником потенциала действия распространяющегося по мембране активно.
Различение сигналов. Закон Вебера-Фехнера. Адаптация сенсорной системы.
Закон Вебера — Фехнера — эмпирический психофизиологический закон, заключающийся в том, что интенсивность ощущения пропорциональна логарифму интенсивности стимула. В ряде экспериментов, начиная с 1834 года, Э. Вебер показал, что новый раздражитель, чтобы отличаться по ощущениям от предыдущего, должен отличаться от исходного на величину, пропорциональную исходному раздражителю. На основе наблюдений Г. Фехнер в 1860 году сформулировал «основной психофизический закон», по которому сила ощущения p пропорциональна логарифму интенсивности раздражителя S: где S — значение интенсивности раздражителя. S0 — нижнее граничное значение интенсивности раздражителя: если S < S0, раздражитель совсем не ощущается. k - константа, зависящая от субъекта ощущения. Так, люстра, в которой 8 лампочек, кажется нам настолько же ярче люстры из 4-х лампочек, насколько люстра из 4-х лампочек ярче люстры из 2-х лампочек. То есть количество лампочек должно увеличиваться в одинаковое число раз, чтобы нам казалось, что прирост яркости постоянен. И наоборот, если абсолютный прирост яркости (разница в яркости «после» и «до») постоянен, то нам будет казаться, что абсолютный прирост уменьшается по мере роста самого значения яркости. Например, если добавить одну лампочку к люстре из двух лампочек, то кажущийся прирост в яркости будет значительным. Если же добавить одну лампочку к люстре из 12 лампочек, то мы практически не заметим прироста яркости. Можно сказать и так: отношение минимального приращения силы раздражителя, впервые вызывающего новые ощущения, к исходной величине раздражителя есть величина постоянная. Закон Вебера — Фехнера можно объяснить тем, что константы скорости химических реакций, проходящих при рецептировании, нелинейно зависят от концентрации химических посредников физических раздражителей или собственно химических раздражителей.
Адаптация сенсорной системы
Сенсорная система обладает способностью приспосабливать свои свойства к условиям среды и потребностям организма. Сенсорная адаптация – это общее свойство сенсорных систем, заключающееся в приспособлении к длительно действующему (фоновому) раздражителю. Существует общая, или глобальная, и локальная, или селективная адаптация. Общая, или глобальная, адаптация проявляется в снижении абсолютной и повышении дифференциальной чувствительности всей сенсорной системы. Субъективно адаптация проявляется в привыкании к действию постоянного раздражителя (например, мы не замечаем непрерывного давления на кожу привычной одежды). Локальная, или селективная, адаптация сводится к снижению чувствительности не всей сенсорной системы, а какой-либо её части, подвергнутой длительному действию стимула.
Адаптационные процессы начинаются на уровне рецепторов, охватывая и все нейронные уровни сенсорной системы. Адаптация слаба только в вестибуло- и проприорецепторах. По скорости данного процесса все рецепторы делятся на быстро- и медленно адаптирующиеся. Первые после развития адаптации практически не посылают в мозг информации о длящемся раздражении. Вторые эту информацию передают в значительно ослабленном виде. Когда действие постоянного раздражителя прекращается, абсолютная чувствительность сенсорной системы восстанавливается. Так, в темноте абсолютная чувствительность зрения резко повышается.
В сенсорной адаптации важную роль играет эфферентная регуляция свойств сенсорной системы. Она осуществляется за счёт нисходящих влияний со стороны более высоких на более низкие её отделы. Происходит как бы перенастройка свойств нейронов на оптимальное восприятие внешних сигналов в изменившихся условиях. Кроме того, состояние разных уровней сенсорной системы контролируется также ретикулярной формацией, включающей их в единую систему, интегрированную с другими отделами мозга и организма в целом. Эфферентные влияния в сенсорных системах чаще всего имеют тормозный характер, т.е. приводят к уменьшению их чувствительности и ограничивают поток афферентных сигналов.
Общее количество эфферентных нервных волокон, приходящих к элементам какого-либо нервного слоя, как правило, во много раз меньше количества его собственных нейронов. Это определяет важную особенность эфферентного контроля в сенсорных системах: его широкий и диффузный характер. Речь идёт об общем снижении чувствительности значительной части нейронного слоя.
Периферический (рецепторный) отдел обонятельной сенсорной системы. Механизм возбуждения обонятельных рецепторов.
Периферический отдел обонятельного анализатора.
Периферическая часть обонятельного анализатора расположена в слизистой оболочке верхнего носового хода и противолежащей части носовой перегородки. Она представлена обонятельными и опорными клетками. Вокруг каждой опорной клетки расположено 9—10 обонятельных. Обонятельные клетки покрыты волосками, которые представляют собой нити длиной 20—30 мкм. Они сгибаются и разгибаются со скоростью 20—50 раз в 1 мин. Внутри волосков расположены фибриллы, которые обычно заходят в утолщение — пуговку, имеющуюся на конце волоска. В теле обонятельной клетки и в ее периферическом отростке расположено большое количество микротрубочек диаметром 0,002 мкм, предполагают, что они осуществляют связь между различными органеллами клетки. Тело обонятельной клетки богато РНК, которая образует возле ядра плотные скопления. После воздействия паров пахучих веществ происходит их разрыхление и частичное исчезновение, что говорит о том, что функция обонятельных клеток сопровождается изменениями в распределении РНК и в ее количестве.
Обонятельная клетка имеет два отростка. Один из них через отверстия продырявленной пластинки решетчатой кости направляется в полость черепа к обонятельным луковицам, в которых возбуждение передается на расположенные там нейроны. Их волокна образуют обонятельные пути, которые подходят к различным отделам ствола мозга. Корковый отдел обонятельного анализатора находится в гиппокамповой извилине и в аммоновом роге.
Второй отросток обонятельной клетки имеет форму палочки шириной 1 мкм, длиной 20—30 мкм и заканчивается обонятельным пузырьком — булавой, диаметр которой 2 мкм. На обонятельном пузырьке расположено 9—16 ресничек.
Механизм возбуждения обонятельных рецепторов.
Обонятельные рецепторы располагаются в слизистой оболочке верхнего отдела полости носа (рис. 331). Они представляют собой особые нервные обонятельные клетки, периферические отростки которых (дендриты) короткие, палочковидные и снабжены чувствительными волосками. Центральные отростки (нейриты) более тонкие, направляются через решетчатую пластинку и продолжаются далее в виде волокон обонятельного нерва. В полости черепа обонятельные нервы входят в обонятельную луковицу, лежащую на основании черепа и составляющую особую долю мозга.
Обонятельные раздражения по волокнам обонятельного тракта передаются в центр обоняния, лежащий рядом с центром вкуса в коре височной доли головного мозга (часть крючковидной извилины морского коня, отчасти аммонов рог).
Проводниковый и корковый отделы обонятельной сенсорной системы.
Проводниковый отдел обонятельного анализатора.
Проводниковый отдел представлен проводящими нервными путями в виде обонятельного нерва, ведущие к обонятельной луковице (образование овальной формы). Проводниковый отдел. Первым нейроном обонятельного анализатора следует считать нейросенсорную или нейрорецепторную клетку. Аксон этой клетки образует синапсы, называемые гломерулами, с главным дендритом митральных клеток обонятельной луковицы, которые представляют второй нейрон. Аксоны митральных клеток обонятельных луковиц образуют обонятельный тракт, который имеет треугольное расширение (обонятельный треугольник) и состоит из нескольких пучков. Волокна обонятельного тракта отдельными пучками идут в передние ядра зрительного бугра.
Корковый отдел обонятельного анализатора.
Ядро обонятельного анализатора у человека расположено в образованиях старой коры, а именно в глубине борозды аммонова рога. Ядра анализатора обоих полушарий связаны друг с другом проводящими путями. К обонятельному анализатору следует отнести и некоторые соседние образования межуточной коры. Прилегающие участки островковой области, лежащие в глубине сильвиевой борозды, по-видимому, имеют то же значение для обоняния, каким обладают проекционно-ассоциативные поля 18 и 19 для зрительной функции.
Есть основания полагать, что к обонятельному анализатору относится и небольшой участок краевой области, расположенной на внутренней поверхности полушария в виде узкой полоски вдоль мозолистого тела. От коркового отдела обонятельного анализатора идут эфферентные пути к нижележащим отделам мозга, в частности к сосковым телам подбугровой области и к уздечке надбугорья. Через эти пути осуществляются корковые рефлексы на обонятельные раздражения.
Периферический (рецепторный) отдел вкусовой сенсорной системы. Механизм возбуждения вкусовых рецепторов.
Периферический отдел вкусового анализатора.
Периферический отдел вкусового анализатора представлен вкусовыми луковицами круглой или овальной формы, которые расположены главным образом в сосочках языка. Различают сосочки желобоватые, листовидные и грибовидные. В меньшем количестве вкусовые луковицы встречаются на мягком небе и задней стенке глотки.Вкусовая луковица имеет овальную форму и состоит из опорных и рецепторных вкусовых клеток. Рецепторные вкусовые клетки усеяны на своем конце микроворсинками, которые называют еще вкусовыми волосками Длина ворсинок – около 2 мкм, диаметр – около 0,2 мкм. Они выходят на поверхность языка через вкусовые поры.
На вкусовой клетке имеется большое число синапсов, которые образуют волокна барабанной струны и языкоглоточного нерва. Волокна барабанной струны (ветвь лицевого нерва) подходят ко всем грибовидным сосочкам, а волокна языкоглоточного нерва – к желобоватым и листовидным. Корковый конец вкусового анализатора находится в гиппокампе, парагиппокамповой извилине и в нижней части заднецентральной извилины.
Вкусовые клетки непрерывно делятся и непрерывно гибнут. Особенно быстро происходит замещение клеток, расположенных в передней части языка, где они лежат более поверхностно. Замена клеток вкусовой почки сопровождается образованием новых синаптических структур.
Механизм возбуждения вкусовых рецепторов.
Как известно, все пищевые вещества в зависимости от вида вкусового ощущения, вызываемого у человека, делятся на четыре основные группы — сладкие, соленые, кислые и горькие. Это объясняется наличием соответственно четырех видов вкусовых рецепторных клеток. Благодаря использованию микроэлектродной техники показано, что одни вкусовые почки реагируют только на горькие вещества, другие — на соленые, третьи — на кислые, четвертые — на сладкие. Рецепторы, чувствительные к разным веществам, неравномерно распределены на поверхности языка. Так, «сладкочувствительные» рецепторы расположены на кончике языка, который особенно чувствителен к сладким веществам, где в основном представлены грибовидные сосочки. Рецепторы, воспринимающие соленый вкус (это, как правило, некоторые соли, в том числе хлористый натрий), разбросаны по всей поверхности языка, т.е. они не имеют «своей» области. Рецепторы, возбуждение которых дает ощущение кислого (главным образом, за счет воздействия свободных водородных ионов), в основном находятся в средней части краев языка (листовидные сосочки). Рецепторы, возбуждение которых дает ощущение горького (при воздействии, например, хинина), находятся, в основном, близ корня языка (желобоватые сосочки).
В основе вкусовой рецепции лежит процесс взаимодействия молекул вкусового вещества с белками-рецепторами, которые представляют собой стереоспецифические участки поверхностной мембраны микроворсинок вкусовых рецепторных клеток. Этапы первичного преобразования химической энергии вкусовых веществ в энергию нервного возбуждения вкусовых рецепторов еще не известны. Можно лишь предположить, что (как и при обонятельной рецепции) активация белка-рецептора должна приводить (за счет каскада промежуточных реакций) к деполяризации рецепторной клетки. Это приведет к выделению медиатора, возбуждающего дендриты вкусовых афферентных нейронов, тела которых расположенных в соответствующих ганглиях головы.
Многие из волокон, идущих от вкусовых рецепторов, отличаются определенной специфичностью, так как отвечают учащением импульсных разрядов лишь на действие соли, кислоты и хинина. Другие волокна реагируют на сахар. Наиболее убедительной считается гипотеза, согласно которой информация о четырех основных вкусовых ощущениях (горьком, сладком, кислом и соленом) кодируется не импульсацией в одиночных волокнах, а разным распределением частоты разрядов в большой группе волокон, по-разному возбуждаемых вкусовым веществом.
В целом можно утверждать, что вкусовое ощущение является мультимодальным, а всевозможные оттенки вкусовых ощущений зависят от множества дополнительных вкусовых и обонятельных раздражений
Проводниковый и корковый отделы вкусовой сенсорной системы.
Проводниковый вкусового анализатора
Вкусовые почки передних 2/3 языка иннервируются от барабанной струны, входящей в состав лицевого нерва, почки задней трети языка, а также мягкого и твердого неба, миндалины – от языкоглоточного нерва; вкусовые почки в области глотки, надгортанника и гортани – от верхнегортанного нерва (часть блуждающего).
Эти нервы – дендриты биполярных нервов нейронов, лежащих в чувствительных ганглиях.
IIнейрон – в ядре одиночного пучка продолговатого мозга.
III нейрон – в таламусе.
Корковый отделы вкусового анализатора.
Отростки нейронов таламуса идут в кору больших полушарий (четвертый нейрон). Центральный, или корковый, отдел вкусового анализатора локализуется в нижней части соматосенсорной зоны коры в области представительства языка. Большая часть нейронов этой области мультимодальна, т. е. реагирует не только на вкусовые, но и на температурные, механические и ноцицептивные раздражители. Для вкусовой сенсорной системы характерно то, что каждая вкусовая почка имеет не только афферентные, но и эфферентные нервные волокна, которые подходят к вкусовым клеткам из ЦНС, благодаря чему обеспечивается включение вкусового анализатора в целостную деятельность организма.
Кожная механорецепция (механизм возбуждения механорецепторов кожи). Кожная терморецепция (механизм возбуждения терморецепторов кожи).
Кожная механорецепция (классификация рецепторов, механизмы их возбуждения). На различных участках кожи человека расположены инкапсулированные механорецепторы, которые иннервируются миелинизированными афферентными волокнами диаметром 5-12 мкм, проводящими импульсы со скоростью 30-70 м/с.
По функции все механорецепторы делятся на
датчики давления стимула,
датчики скорости его воздействия
датчики ускорения.
В коже имеется также множество так называемых свободных нервных окончаний, на которых отсутствуют корпускулярные структуры. Афферентные волокна свободных нервных окончаний — не-миелинизированные аксоны со скоростью проведения 1 м/с. Геометрия свободных нервных окончаний весьма разнообразна. Свободные нервные окончания реагируют импульсом не более, чем на один уровень интенсивности — пороговый. Поэтому они считаются пороговыми датчиками, отмечающими наличие стимула в определенном месте кожи. Свободные нервные окончания принимают участие в передаче информации о слабых (околопороговых) движущихся по коже стимулах и имеют отношение к ощущению щекотки.
Ощущение давления или прикосновения (тактильные ощущения) можно вызвать только в определенных точках кожи, которые называются чувствительными тактильными точками. Большое число тактильных точек находится на коже кончиков пальцев и на губах. На коже, покрывающей плечи, бедра и спину тактильные точки встречаются редко, для появления тактильного ощущения на внутренней стороне руки достаточно вдавить кожу на глубину 0,01 мм (10 мкм). Пороги тактильных ощущений на кончиках пальцев значительно ниже, чем на остальной поверхности кисти. При помощи ножек циркуля с тупыми концами легко определяются пространственные пороги различения, то есть минимальные расстояния между двумя тактильными стимулами, при которых оказывается возможным воспринять их как раздельные. Одновременное касание кожи кончиками циркуля демонстрирует пороги одновременного пространственного различения. Эти пороги рассматриваются в качестве меры пространственного различения кожей тактильных стимулов в исследуемой части тела.
Из всех кожных механорецепторов наиболее изучены тельца Пачини — детекторы коротких механических воздействий. Многократное частое раздражение этих рецепторов вызывает ощущение вибрации. Капсула тельца Пачини состоит их множества одинаковых слоев, вложенных друг в друга наподобие "матрешки". В сердцевине "матрешки" находится нервное окончание. Эта многослойная конструкция работает как высокочастотный фильтр: при кратковременном воздействии капсула деформируется и действует на нервное окончание. Если действие механического раздражителя продолжается, то происходит изменение расстояний между слоями капсулы, которое компенсирует постоянное давление на тельце Пачини и приводит к отсутствию деформации нервного волокна.
Капсула тельца Пачини деформируется при воздействии механического раздражителя и действует на нервное окончание. В нервном окончании возникает генераторный потенциал. Поскольку капсула работает как высокочастотный фильтр, то она не реагирует на стимуляцию постоянным давлением. В самом возбудительном процессе нейрона капсула не участвует. Она только меняет характер раздражения. При многократном раздражении генераторные потенциалы телец Пачини снижаются.
Потенциалы действия нервных волокон, отходящих от телец Пачини. следуют частоте раздражения до 200-400 Гц, в интервале 400- 500 Гц соответствие импульсации частоте стимула нарушается, выше 500 Гц они перестают реагировать. Люди при одинаковом раздражении кожи ладони отмечают два вида ошушений — вибрации и трепетания. Для возникновения ощущения вибрации необходима более высокая частота и меньшая амплитуда, чем для ощущения трепетания, которое появляется при более низких частотах, но большей амплитуде колебаний. Трепетание исчезает при поверхностной анестезии, так же как разряды рецепторов, реагирующих на низкие частоты вибрации. Местная анестезия поверхностных слоев кожи не уничтожает вибрационную чувствительность и ответы высокочастотных рецепторов. Условием появления ощущения вибрации в коже является одновременное вовлечение в реакцию нескольких телец Пачини, поскольку именно в этом случае разряд в афферентных волокнах становится эффективным стимулом.
Механизмы возбуждения кожных рецепторов. Механический стимул приводит к деформации мембраны рецептора. В результате этого электрическое сопротивление мембраны уменьшается, т.е. увеличивается её проницаемость для ионов. Через мембрану рецептора начинает течь ионный ток, приводящий к генерации рецепторного потенциала. При достижении рецепторным потенциалом критического уровня деполяризации генерируются импульсы, распространяющиеся по волокну в ЦНС.
Механизм возбуждения:
Стимул
Деформация мембраны рецептора
Уменьшается электрическая сопротивляемость мембраны
Проницаемость для натрия
Генерация РП
Суммация РП
Возникновения ГП
Восприятие прикосновения обеспечивают:
Тельца Мейснера (быстро-адаптирующиеся рецепторы)
Свободные окончания нервных волокон (вдоль мелких сосудов)
Нервные волокна, оплетающие волосяную сумку
Чувство давления – диски Меркеля
образуют рецептивные поля в глубоких слоях кожи и слизистой.
Реагируют на прогибание эпидермиса под действием механического импульса.
Медленно адаптируются
Чувство вибрации – тельца Пачини (детекторы коротких механических воздействий)
Участок кожи не покрытые волосами
Очень быстро-адаптирующиеся рецепторы
Реагирует на 40-1000 гЦ
Максимальная чувствительность – 300 гЦ (более 500 гЦ снижается)
Для появления ощущения вибрации – вовлекаются несколько тел
Щекотание – свободные некапсулированные нервные окончания, расположенные в поверхностных слоях кожи (появление и передвижение стимула)
Кожная терморецепция (классификация рецепторов, механизмы их возбуждения).
Роль – поддержание температуры тела на постоянном уровне.
Виды рецепторов:
Первично-чувствующие (контактные термо-рецепторы)
Свободные нервные окончания
Специфические (температурные)
Неспецифические
Терморецепция – восприятие и анализ информации с целью обеспечения терморегуляции, доставляя информацию к центру темпуратуры (гипоталамус)
Холодовые рецепторы (миелинизированные, больше)
Колбы Краузе
Поверхностный слоя
Порог восприятия температуры меньше (абсолютный и дифференциальный), а чувствительность больше
Более широкий диапазон восприятия температурных воздействий
10-41 градусов
Тепловые рецепторы (немиелинизированные, меньше)
Тельца Руффини
Глубокий слой
Порог восприятия температуры больше, чувствительность – меньше
Более узкий диапазон восприятия температурных воздействий
20-50 градусов
Механизм возбуждения:
Изменение температуры
Активация ионных каналов
Суммация РП
Возникновение ПД
Статитическая импульсация – постоянная частота ПД. Наблюдается при температуре 18-27 градусов (температурный комфорт).
Динамическая импульсация – изменение количества импульсации при изменении температуры окружающего мира.
Специальные рецепторы тепла и холода выполняют две основные функции — они отвечают на изменения температуры окружающей среды и участвуют в регуляции температуры тела. Рецепторы кожи, чувствительные к холоду и теплу (точки тепла и холода), расположены на разных участках тела. Плотность и общее число температурных точек меньше, чем число тактильных точек во всех областях тела человека. Точек холода на коже значительно больше, чем точек тепла., например, на поверхности кисти руки имеется по 1-5 точек холода на каждом квадратном сантиметре и лишь в среднем по 0,4 точки тепла.
Низкая, по сравнению с плотностью механорецепторов, плотность точек холода и, особенно, тепла свидетельствует, что одновременные пространственные пороги для температурных стимулов относительно велики. Пространственные пороги для холодовых стимулов ниже, чем для тепловых.
Специфические терморецепторы (рецепторы холода и тепла) предположительно представляют собой свободные нервные окончания, которые несут также механорецепторные функции.У человека рецепторы холода располагаются в эпидермисе и непосредственно под ним, а рецепторы тепла — преимущественно в верхнем и среднем слоях собственно кожи. Рецепторы холода связаны с тонкими миелинизи-рованными волокнами, а рецепторы тепла — с немиелинизированны-ми волокнами. Специфические терморецепторы характеризуются рядом общих показателей. Так, при постоянной температуре кожи они тонически разряжаются с частотой, зависящей от температуры —статическая реакция. В случае изменения температуры кожи они увеличивают или уменьшают частоту разряда —динамическая реакция. Специфические терморецепторы нечувствительны к нетепловым стимулам, а пороги их импульсной реакции сравнимы с порогами появления ощущений при тепловой стимуляции кожи.
Ощущения, появляющиеся при изменении температуры кожи, определяются: 1)исходной температурой кожи, 2) скоростью изменения температуры кожи, 3) площадью кожи, на которую действует стимул.
Исходная температура влияет на пороги ощущения тепла и холода. При низких температурах, например при 20°С, порог для появления ощущения тепла высокий, а для появления ощущения холода — низкий. Если исходную температуру кожи повысить, то тепловые пороги уменьшатся, а холодовые увеличатся. При этом следует иметь в виду, что в зависимости от условий стимуляции одна и та же температура кожи может вызывать и ощущение тепла, и ощущение холода.
Скорость изменения температуры, если она превышает 0,1°С с (6°С мин), почти не оказывает влияния на пороги ощущения тепла и холода. При снижении скорости изменения температуры оба порога монотонно возрастают.
Площадь кожи, на которой меняется температура, отражается на порогах ощущений при охлаждении и нагревании, соответственно, больше в случае малых площадей, чем при больших. При подобном надпороговым изменении температуры кожи интенсивность ощущений возрастает с увеличением стимулируемой площади. Следовательно, и в околопороговом, и в надпороговом диапазонах происходит пространственная суммация импульсных реакций терморецепторов в нервных центрах.
Мышечно-сухожильная и суставная проприорецепция: мышечные веретена, их характеристика и механизм возбуждения; сухожильные рецепторы Гольджи, их характеристика и механизм возбуждения.
Имеется 3 вида рецепторов: мышечные веретена, сухожильные рецепторы Гольджи и суставные рецепторы.
Мышечное веретено: одето капсулой и расположено в толще мышцы. Внутри капсулы находится пучок интрафузальных мышечных волокон. Веретена расположены параллельно внешним по отношению к капсуле экстрафузальным волокнам, поэтому при растяжении мышцы нагрузка на веретена увеличивается, а при сокращении – уменьшается. В расслабленной мышце импульсация, идущая от веретен, невелика, но они реагируют повышением частоты разрядов на удлинение мышцы. Таким образом, веретена дают мозгу информацию о длине мышцы и ее изменениях. Импульсация, идущая от веретен, в спинном мозге возбуждает мотонейроны своей мышцы и тормозит мотонейроны мышцы-антагониста, а также возбуждает мотонейроны сгибателей и тормозит мотонейроны разгибателей.
Сухожильные рецепторы Гольджи: располагаются в месте перехода мышечных волокон в сухожилия. Отдельный нервный аксон несет афферентные импульсы в спинной мозг и называется аксоном Ib. Он начинается в виде веточек, проходящих между коллагеновыми волокнами сухожилия. Когда мышечные волокна сокращаются, коллагеновые волокна натягиваются и сжимают нервные веточки, которые начинают импульсировать. Таким образом, в результате последовательного крепления сухожильных органов к мышечным волокнам они возбуждаются при укорочении возбужденной мышцы. - суставные рецепторы.
Тельца Руффини: находятся в капсуле сустава и воспринимают направление и скорость изменения межзвенного угла. Частота их импульсации возрастает с увеличением скорости изменения суставного угла.
Тельца Пачини посылают в ЦНС информацию о положении отдельных частей тела в пространстве и относительно друг друга. Эти рецепторы посылают в ЦНС информацию о значениях межзвенных углов, то есть о положении сустава. Их импульсация продолжается в течение всего периода сохранения межзвенного угла, и она тем больше, чем больше изменения угла.
Проводниковый и корковый отделы соматосенсорной системы: лемнисковый и спиноталамический пути проведения, их характеристика. Сенсорный гомункулюс.
Проводниковый отдел соматовисцеральной сенсорной системы
Основные функции проводникового отдела — кодирование информации о стимуле и проведение её в кору больших полушарий. Он включает чувствительные волокна черепных и спинномозговых нервов, внутримозговые проводящие пути, подкорковые центры первичного анализа в стволе.
Начинается проведение нервного импульса с образованием ПД. Натрий поступает в клетку нервного волокна и возникает электрические ионные токи. Их возникновение связано с разными зарядами по обе стороны мембраны нервного волокна. Установлено, что распространение нервного импульса по мякотному волокну быстрее, чем по безмякотному. Мякотное волокно покрыто миелином, который является электроизолятором.
Корковый отдел соматовисцеральной сенсорной системы. Понятие о сенсорном гомункулусе.
Корковый (центральный) отдел сенсорной системы расположен в коре больших полушарий, причём у каждого имеется своё представительство. Корковый отдел состоит из ядра или ядерной зоны и периферической зоны (зоны рассеянных элементов). Установлено, что ядерная зона выполняет наиболее сложные задачи анализа и синтеза поступающей информации. В зоне рассеянных элементов осуществляется элементарный анализ и синтез. Ядерная зона расположена более локально, а периферическая представлена в виде разбросанных отдельных элементов по всей доле мозга. Известный нейропсихолог Лурия выделил 3 основных зоны коркового отдела сенсорных систем: 1. Первичная (проекционная) зона обеспечивает мелкий синтез поступающих сигналов, т.е. она определяет узкоспецифичные характеристики (стимулы). 2. Вторичная (проекционно-ассоциативная), она как бы надстраивается над первичной и осуществляет более сложные формы анализа и синтеза. Она отвечает за узнавание. 3. Ассоциативная, она надстраивается над вторичной, самая молодая и выполняет наиболее сложные функции. На основании информации, поступающей из первичных и вторичных зон, третичная зона вырабатывает цели и задачи поведения.
Передача и переработка соматосенсорной информации.
2 основных пути (тракта): лемнисковый и спинноталамический.
Лемнисковый путь.
Отличительная особенность – в быстрой передаче в мозг наиболее точной информации, дифференцированной по силе и месту воздействия. Передает сигналы о прикосновении к коже, давлении на нее и движениях в суставах.
На всех уровнях состоит из относительно толстых, быстропроводящих миелинизированных нервных волокон. Первые нейроны находятся в СМ узле, их аксоны в составе задних столбов восходят к тонкому (ядро Голля) и клиновидному (ядро Бурдаха) ядрам ПрМ, где сигналы передаются на 2-е нейроны лемнискового пути.
В ПрМ в тонком ядре сосредоточены в основном 2-е нейроны тактильной чувствительности, а в клиновидном – 2-е нейроны проприоцептивной чувствительности. Аксоны этих нейронов образуют медиальную петлю и после перекреста на уровне олив направляются в специфические ядра таламуса – вентробазальный ядерный комплекс. В этих ядрах концентрируются 3-и нейроны лемнискового пути. Их аксоны направляются в соматосенсорную зону КБМ.
Удаление соматосенсорной зоны коры приводит к нарушению способности локализовать тактильные нарушения, а электростимуляция вызывает ощущение прикосновения, вибрации и зуда.
В целом роль коры в интегральной оценке соматосенсорных сигналов, во включении их в сферу сознания, полисенсорный синтез и в сенсорное обеспечение выработки новых двигательных навыков.
Спинноталамический путь.
Более медленная передача афферентных сигналов, со значительно менее четко дифференцированной информацией о разных свойствах раздражителя и с менее четкой топографической локализацией. Служит для передачи температурной, всей болевой и в значительной мере – тактильной чувствительности.
1-е нейроны расположены в спинномозговом узле, откуда посылают в СМ медленнопроводящие немиелинизированные нервные волокна. Эти нейроны имеют большие рецептивные поля. 2-е нейроны локализуются в сером веществе СМ, а их аксоны в составе восходящего спинно-таламического пути направляются после перекреста на спинальном уровне в вентробазальный ядерный комплекс таламуса (дифференцированные проекции), а также в вентральные неспецифические ядра таламуса, внутреннее коленчатое тело, ядра ствола мозга и гипоталамус, где локализуются 3-и нейроны. 3-и нейроны лишь частично дают проекци и в соматосенсорную зону коры.
Вместе с тем, от специфических ядер таламуса импульсация поступает в соматосенсорную кору в области постцентральной извилины и глубине сильвиевой борозды. Окончательное отношение к боли возникает с участием нейронов лобной доли коры. Одновременно поток импульсации от ноцицепторов на уровнях продолговатого и среднего мозга отходит по коллатералям к неспецифическим ядрам таламуса, от них ко всем участкам коры (диффузная активация), а также достигает нейронов лимбической системы, благодаря чему приобретается эмоциональная окраска. Т.о., эта система играет важную роль в организации генерализованных ответов на действие болевых, температурных и тактильных раздражителей.
Сенсорный (соматосенсорный) гомункулус
Кортикальные клетки, чьи рецептивные поля располагаются на поверхности тела рядом, группируются вместе и в коре. В результате в первичной соматосенсорной коре образуется представительство контрлатеральной поверхности тела, которое может быть картировано с помощью микроэлектродной техники. Карта поверхности туловища и лица человека, находящаяся на постцентралыюй извилине, получила название "сенсорный гомункулус". Изображения на этой карте непропорциональны, поскольку наибольший объем нервной ткани относится к областям тела с особенно плотной иннервацией - область вокруг рта, большой палец руки и остальные пальцы. Такое представительство иногда называют соматосенсорным гомункулусом.
Впервые это явление было обнаружено канадским нейрохирургом Уайлдером Пэнфилдом в ходе операций на головном мозге при местной анестезии. Чтобы убедиться, что те или иные иссечения мозга не приведут к катастрофическим последствиям, Пенфилд стимулировал кору в различных точках и спрашивал пациента, что он чувствует. Поскольку размеры РП кортикальных клеток варьируют так сильно, получившаяся карта чрезвычайно неизоморфна.
Интерорецепторы, их характеристика. Проводящие пути и центры висцеральной сенсорной системы.
Висцерорецепция – это восприятие висцерорецепторами изменений условий внутренней среды и состояния внутренних органов, передача информации в ЦНС и восприятие ее сенсорными нейронами коры больших полушарий.
Рецепторы висцеральной системы воспринимают разнообразные изменения внутренней среды организма, посылают информацию в ЦНС, участвуя таким образом в работе внутренних органов и поддержании гомеостаза в организме (химического состава, осмотического давления, температуры внутренней среды, давления крови в сосудах, давления и наполнения внутренних полых органов).
Периферический отдел висцеральной системы представлен соответствующими рецепторами. Висцероцепторы находятся во внутренних органах, тканях, сосудах (механо-, хемо-, термо- и осморецепторы).
Механорецепторы (баро- и волюморецепторы) расположены в сосудах, легких, ЖКТ и др. Для них адекватным стимулом является растяжение, деформация стенок органов.
Хеморецепторы находятся в аортальных и каротидных клубочках, в слизистых оболочках пищеварительного тракта, органов дыхания, в серозных оболочках, в головном мозгу.
Осморецепторы локализованы в аортальном и каротидном синусах, в интерстициальной ткани вблизи капилляров, внутри сосудов артериального русла, в печени и др. органах, а также в гипоталамусе (нейроны – осмометры). Часть осморецепторов является механорецепторами, часть – хеморецепторами (например, Na+ -рецепторы, гликорецепторы).
Терморецепторы находятся в слизистой оболочке пищеварительного тракта, органов дыхания, мочевого пузыря, в серозных оболочках, в стенках артерий и вен, в каротидном синусе, а также в ядрах гипоталамуса.
Все рецепторы висцеральной системы являются первично-чувствующими (за исключением вторично-чувствующих хеморецепторов каротидного клубочка).
Проводниковый отдел представлен в основном чувствительными волокнами вегетативных и соматических нервов, частично-смешанными нервами, в состав которых входят вегетативные волокна.
Чувствительные нейроны находятся в спинальных ганглиях или ганглиях черепно-мозговых нервов, промежуточные – в спинном мозге или в стволе мозга. Восходящие пути от них поступают к заднемедиальным ядрам таламуса и от него к лимбической системе. Проекции висцеральных нервов обнаружены также в мозжечке.
Корковый отдел локализуется в соматосенсорной области и в орбитальной части коры большого мозга. В коре замыкаются натуральные вегетативные условные рефлексы, являющиеся важным приспособительным ответом организма в различных условиях жизнедеятельности. Восприятие некоторых интероцептивных стимулов может сопровождаться возникновением четких локализованных ощущений:например, при растяжении стенок желудка, мочевого пузыря или прямой кишки. Но висцеральная импульсация от сердца, печени,почек обычно не вызывает ощущений, что является сигналом благополучного состояния организма.
Физиологическая роль боли. Теории происхождения боли.
Болевая (ноцицептивная) чувствительность – имеет большое значение для выживания организма, так как:
Сигнализует об опасности при действии различных агентов
Симптом многих заболеваний
Важный показатель диагностики
Ноцицептивная чувствительность — это восприятие стимулов, вызывающих в организме ощущение боли. Ощущение боли коренным. образом отличается от других сенсорных модальностей содержанием информации, которую она в себе заключает. Боль информирует организм об опасности, о нарушениях в процессах, протекающих в различных органах и тканях, поскольку вызывается повреждающими, вредными для организма стимулами. Трудно переоценить значение этой информации для клиники, так как именно болевое проявление действия вредных факторов чаще всего приводит пациента к врачу. Более того, ощущение боли, при всей его субъективности, оказывается часто решающим для диагноза заболевания.
Теории возникновения боли.
Теория специфичности утверждает, что боль представляет собой отдельную сенсорную систему, в которой любой повреждающий стимул активирует специальные болевые рецепторы (ноцицепторы), передающие болевой импульс по специальным нервным путям в спинной мозг и в болевые центры головного мозга, вызывая ответную защитную реакцию, направленную на удаление от раздражителя.
Теория интенсивности утверждает, что ощущение боли возникает при раздражении любого рецептора избыточным стимулом (шум, свет).
Теория воротного контроля Поток болевой импульсации с периферии идет в задний рог спинного мозга по большим миелинизированным (А-дельта) и малым немиелинизированным (С-волокнам) нервным волокнам. Оба типа волокон образуют синапсы с нейронами второго порядка (Т) ("передача/проекция"). Когда Т-нейроны активированы, они поставляют ноцицептивную информацию в мозг.
Периферические нервные волокна также образуют синапсы с интернейронами желатинозной сустанции (ЖС), которые при стимуляции угнетают Т-нейроны. А-дельта волокна стимулируют, а С-волокна угнетают интернейроны ЖС, соответственно снижая и повышая центральную передачу ноцицептивных входящих сигналов.
Кроме того, стимуляция интернейронов ЖС на подавление активности Т-нейронов происходит через нисходящие пути, начинающиеся в центральной нервной системе (это происходит при активации различными факторами). Баланс между возбуждающими и угнетающими сигналами определяет степень передачи ноцицептивной информации в головной мозг («+» - возбуждающий сигнал; «-» - угнетающий сигнал) (рис, 2.7.2.).
А-дельта волокна |
 С-вюлокна
С-вюлокна
Классификация физиологической боли. Отраженная и проецированная боль, механизмы их развития.

Существуют и особые виды боли — проецируемая и отраженная. В качестве примера причины проецируемой боли можно привести резкий удар по локтевому нерву, проходящему в области локтя у самой поверхности. Подобный удар вызывает очень неприятное, трудноописываемое ощущение, распространяющееся на те участи руки, которые обслуживаются этим нервом — на участок от локтя до кисти и на саму кисть. В принципе, проецируемые ощущения могут возникать в рамках любой сенсорной модальности, причем, многие длительные проецируемые боли, в отличие от боли от удара по локтевому нерву, имеют клиническое значение. Механизм их возникновения объясняется следующим. Сигналы, вызываемые ненормальной стимуляцией афферентного волокна, передаются обычно в мозг по латеральному спинно-таламическому тракту, порождая там ощущения, относимые к области, которую иннервирует данное афферентное волокно. Одна из распространенных причин проецируемых болей — это пережатие спинальных нервов в местах их вхождения в спинной мозг в результате повреждения межпозвонковых хрящевых дисков. Афферентные импульсы в ноцицептивных волокнах при такой патологии вызывают болевые ощущения, которые проецируются в область, связанную с травмируемым спинальным нервом.
Отраженная боль отличается от проецирумой боли тем, что она вызывается не прямой стимуляцией нервных волокон, а раздражением каких-либо рецептивных окончаний. Отраженными болями называются болевые ощущения не во внутренних органах, от которых поступают болевые сигналы, а в расположенных на некотором расстоянии от больного органа поверхностных областях. Хорошо известный пример отраженной боли — боль, возникающая в сердце, но ощущаемая в плече и в узкой полоске на медиальной поверхности руки. Так как взаимоотношения между отдельными участками кожи (дерматомами) и внутренними органами хорошо известны, подобные отраженные боли играют большую роль в диагностике различных заболеваний. Механизм возникновения отраженной боли сводится к следующему. Некоторые кожные болевые афференты и болевые эфференты, идущие от внутренних органов, при вхождении в один и тот же сегмент спинного мозга могут конвергировать на один и тот же нейрон. Возбуждение таких клеток на периферии интерпретируется человеком как боль.
Система подавления боли (антиноцицептивная система). Локальный и нисходящий контроль боли.
Характеристика антиноцицептивной системы. Функция антиноцицептивной системы заключается в контроле над активностью ноцицептивной системы и предотвращении ее перевозбуждения. Ограничительная функция проявляется увеличением тормозного влияния антиноцицептивной системы на ноцицептивную систему в ответ на нарастающий по силе болевой стимул. Первый уровень представлен комплексом структур среднего, продолговатого и спинного мозга, к которым относятся околоводопроводное серое вещество, ядра шва и ретикулярной формации, а также желатинозная субстанция спинного мозга. Структуры этого уровня объединяются в морфофункциональную «систему нисходящего тормозного контроля». Медиаторами являются серотонин и опиоиды. Второй уровень представлен гипоталамусом, который: 1) оказывает нисходящее тормозное влияние на ноцицептивные структуры спинного мозга; 2) активизирует систему «нисходящего тормозного контроля», т. е. первый уровень антиноцицептивной системы; 3) тормозит таламические ноцицептивные нейроны. Медиаторами этого уровня являются катехоламины, адренергические вещества и опиоиды. Третьим уровнем является кора больших полушарий, а именно II соматотропная зона. Этому уровню отводится ведущая роль в формировании активности других уровней антиноцицептивной системы формирование адекватных реакций на повреждающие факторы.
Механизм деятельности антиноцицептивной системы. Антиноцицептивная система оказывает свое действие посредством: 1) эндогенных опиоидных веществ: эндорфинов, энкефалинов, и динорфинов. Эти вещества, связываются с опиоидными рецепторами, имеющимися во многих тканях организма, особенно в ЦНС. 2) В механизме регуляции болевой чувствительности участвуют и неопиоидные пептиды: нейротензин, ангиотензин II, кальцитонин, бомбезин, холецистокинин, которые оказывают также тормозной эффект на проведение болевой импульсации. 3) В купировании определенных видов боли участвуют и непептидные вещества: серотонин, катехоламины.
Строение и функции вестибулярного аппарата. Характеристика рецепторов вестибулярного аппарата, механизм вестибулорецепии.
Функции вестибулярного анализатора. Организация вестибулярного аппарата.
Функции вестибулярного анализатора:
Пространственная ориентировка
Поддержание позы (равновесия)
Стабилизация взгляда (динамическая острота зрения)
Перераспределение тонуса скелетных мышц
Восприятие информации о ускорении, замедлении, изменении положения головы
!!Рецепторы вестибулярной системы не возбуждаются при:
Равномерном движении
В состоянии покоя
Строение вестибулярного анализатора
Периферический отдел – костный лабиринт пирамиды височной кости. Представлен:
3 полукружных канальца (взаимно перпендикулярное направление)
Отолитовый орган (преддверный орган) – мешочек и маточка
Перепончатый лабиринт (мешочек, маточка, полукружные канальца) – состоят из тонких перепонок, которые образуют замкнутые трубки (внутри эндолимфа)
Перилимфа – между костным и перепончатым лабиринтом
Мешочки преддверия
Функции- восприятие линейных ускорений, наклона головы или всего тела; силы тяжести.
Отолитовый аппарат – располагаются на макулах (возвышение в мешочках), это скопление рецепторных волосков и опорных клеток.
Волоски (цилии) – располагаются на поверхности рецепторных клеток и обращены в полость (просвет) перепончатого лабиринта. Различают:
Киноцилий – самый длинный, 1
Стереоцилий – тонкие, 60-80 шт
Волоски погружены в отолитовую мембрану (отолит – кристаллы карбоната кальция)
При наклоне за счёт силы тяжести отолит смещает мембрану относительно сенсорного эпителия и цилий, они погружаются в неё и сгибаются благодаря скольжению мембраны вдоль них.
Полукружные канальца
Функции – восприятие угловых ускорений
Рецепторные клетки сконцентрированы в ампулах (расширение в конце каждого полукружного канальца) в виде Крист
Купула – желеобразное образование, которое вдается в эндолимфу (в нем нет отолитов; удельный вес равен удельному весу эндолимфы)
Угловое ускорение воспринимается в силу иннервации эндолимфы
Купула сгибается, погружённые в неё Цилии рецепторных клеток оказываются под действием смещения и происходит их адекватное раздражение.
Вестибулярная сенсорная система состоит из следующих отделов:периферический отдел включает два образования, содержащие механорецепторы вестибулярной системы — преддверие (мешочек и маточка) и полукружные каналы;
проводниковый отдел начинается от рецепторов волокнами биполярной клетки (первого нейрона) вестибулярного узла, расположенного в височной кости, другие отростки этих нейронов образуют вестибулярный нерв и вместе со слуховым нервом в составе 8-ой пары черепно-мозговых нервов входят в продолговатый мозг; в вестибулярных ядрах продолговатого мозга находятся вторые нейроны, импульсы от которых поступают к третьим нейронам в таламусе (промежуточный мозг);
корковый отдел представляют четвертые нейроны, часть которых представлена в проекционном (первичном) поле вестибулярной системы в височной области коры, а другая часть — находится в непосредственной близости к пирамидным нейронам моторной области коры и в постцентральной извилине. Точная локализация коркового отдела вестибулярной сенсорной системы у человека в настоящее время не установлена.
Механизм возбуждения вестибулорецепторов.
Преддверие – при прямом положении тела; утрикулюс – горизонтально; саккулюс-вертик
Стимул (наклон)
Смещение мешочков
Сгибание цилия и погружение в отолитовую мембрану
Сгибание киноцилия – решающая роль в генерации импульса
В сторону киноцилия
Усиление импульса
Открытие механоупр. К-каналов
Деполяризация
Генерация РП
Выход кальция и АХ
Постсинаптическая мембрана вестибулярного нерва
Генеративный потенциал
Наоборот, если от киноцилия – уменьшение импульса и гиперполяризация
!!!Волокна вестибулярного нерва – находятся в постоянной активности, смещение киноцилия ведёт к изменению импульсации (частоты раздражения)
Волокна вестибулярного нерва (отростки биполярных нейронов) направляются в продолговатый мозг. Импульсы, приходящие по этим волокнам, активируют нейроны бульбарного вестибулярного комплекса, в состав которого входят ядра: преддверное верхнее, или Бехтерева, преддверное латеральное, или Дейтерса, Швальбе и Роллера. Отсюда сигналы направляются во многие отделы ЦНС: спинной мозг, мозжечок, глазодвигательные ядра, кору большого мозга, ретикулярную формацию и ганглии автономной нервной системы.
Электрические явления в вестибулярной системе.
Даже в полном покое в вестибулярном нерве регистрируется спонтанная импульсация. Частота разрядов в нерве повышается при поворотах головы в одну сторону и тормозится при поворотах в другую (детекция направления движения). Реже частота разрядов повышается или, наоборот, тормозится при любом движении. У 2/з волокон обнаруживают эффект адаптации (уменьшение частоты разрядов) во время длящегося действия углового ускорения. Нейроны вестибулярных ядер обладают способностью реагировать и на изменение положения конечностей, повороты тела, сигналы от внутренних органов, т. е. осуществлять синтез информации, поступающей из разных источников.
Характеристика рецепторов вестибулярного аппарата, механизм вестибулорецепии.
Они также снабжены волосками. При движении эндолимфы (во время угловых ускорений), когда волоски сгибаются в одну сторону, волосковые клетки возбуждаются, а при противоположно направленном движении - тормозятся. В волосковых клетках и преддверия, и ампулы при их сгибании генерируется рецепторный потенциал, который через синапсы (посредством выделения ацетилхолина) передает сигналы о раздражении волосковых клеток окончаниям волокон вестибулярного нерва. Возбуждение волосковых клеток преддверия происходит вследствие скольжения отолитовой мембраны по волоскам, т.е. их сгибания.
Проводниковый и корковый отделы вестибулярной сенсорной системы. Вестибулярные рефлексы, их характеристика. Нистагм глаз.
Проводниковый и корковый отделы вестибулярного анализатора.
Автоматический контроль
I нейрон – генерация ПД , дендриты афферентных биполярных клеток вестибулярного
ганглия (аксон идет к продолговатому мозгу)
II нейрон – вестибулярные ядра продолговатого мозга (ядра Бехтерева, Дейтриса,
Швальбе, Роллера)
III нейрон :
Мозжечок
Ядра глазодвигательного нерва
Tr.vestibulospinalis к мотонейронам мышц разгибателей
К ретикулярной формации
Гипоталамусу
Таламическими ядрами
Происходит без участия сознания (поддержание позы и равновесия, рефлексы)
Сознательный анализ
Прямой путь – таламокортикальный (осознание положения тела в пространстве, восприятие перемещений)
Первый и второй нейрон – как и в авт.контроле
III нейрон – таламус (дорсо-медиальная часть вентрального постлатерального ядра)
IV нейрон – задняя постцентральная извилина
Непрямой путь – вестибуло-церебелло-таламический (подержание тонуса мыщц, который связан с оценкой позы тела и схемы тела)
III нейрон – мозжечок
IV нейрон – таламус (медиальная часть вентрального ядра)
V нейрон – моторная кора спереди от нижней центральной извилины
проводниковый отдел. К рецепторам подходят периферическиеволокна биполярных нейронов вестибулярного ганглия, расположенного во вутреннем слуховом проходе (первый нейрон). Аксоны этих нейронов в составе вестибулярного нерва направляются квестибулярым ядрам продолговатогомозга (второй нейрон). Вестибулярные ядра продолговатого мозга (верхнее—ядро Бехтерева,медиальное—ядро Швальбе,латеральное—ядро Дейтерсаинижнее—ядро Роллера) получают дополнительную информацию по аф- ферентным нейронам от проприорецепторов; мышц или от суставных сочленений шейного отдела позвоночника. Эти ядра, где расположен второй нейрон вестибулярного анализатора, тесносвязаны с различными отделами центральной нервной системы.Благодаря этому обеспечиваются контроль и управление эффекторными реакциями соматического, вегетативного и сенсорного характера.
третий нейронрасположен вядрах зрительного бугра, откуда возбуждение направляется в кору большого полушария.
Центральный отдел вестибулярного анализатора локализуется ввисочной области коры большого мозга, несколько кпереди от слуховой проекционной зоны (21—22-е поля по Бродману, четвертый нейрон).
Нервные волокна, выходящие из вестибулярных ядер, образуют связи с другими отделами центральной нервной важнейшими из них являются следующие: вестибулоспинальные, вестибуловегетативныеивестибулоглазодвигательные. Эти связи обеспечивают контроль и управление различными двигательными реакциями, а также являются основой для рефлексов обеспечивающих равновесие.
а. Вестибулоспинальный тракт, волокна которого в конечном итоге оказывают влияние в основном наγ-мотонейроны мышц-разгибателей, хотя часть волокон оканчивается и наα-мотонейронах.
б. Связи с мотонейронамишейного отдела спинного мозга, входящие главным образом в вестибулоспинальный тракт.
в.Связи с ядрамиглазодвигательного нерва, которые опосредуют движения глаз, вызываемые вестибулярной активностью; эти волокна проходят в составе медиального продольного пучка. Функцию статолитового аппарата можно тестировать, наблюдая глазодвигательные реакции при наклонах головы.
г. Тракты, направляющиеся ввестибулярные ядра противоположной стороны мозга, благодаря которым афферентация с обеих сторон тела может обрабатываться совместно.
д. Связи сретикулярной формацией, посредством которых обеспечивается воздействие на ретикулоспинальный тракт, являющийся еще одним (полисинаптическим) путем кα- иβ-мотонейронам.
е. Тракты, проходящие через таламус впостцентральную извилину коры головного мозга, обеспечивающие сознательную обработку вестибулярной информации и таким образом сознательную ориентацию в пространстве.
ж.Волокна, направляющиеся вгипоталамус, которые в основном участвуют в возникновении кинетозов (укачивания).
з. Связи смозжечком, в особенности сarchicerebellum( вторичные вестибулярные афференты).мозжечок, может получать некоторые первичные вестибулярные афференты (так называемый прямой сенсорный мозжечковый путь). И первичные, и вторичные вестибулярные афференты у млекопитающих оканчиваются в мозжечке мшистыми волокнами на клетках-зернах флокку-лонодулярной доли (относящейся кarchicerebellum) и частично клеткахuvulaиparaflocculus(paleocerebellum). Клетки-зерна оказывают возбуждающее действие на клетки Пуркине в этих областях, а аксоны последних проецируются опять-таки в вестибулярные ядра. Такая цепь осуществляет тонкую регулировку вестибулярных рефлексов. При нарушениях функции мозжечка вследствие различных заболеваний эти рефлексы утрачивают тормозной компонент, что проявляется в возникновении таких симптомов, как усиленный или спонтанно возникающий нистагм, утрата равновесия, проявляющаяся в неустойчивой походке, избыточной амплитуде движений, особенно при ходьбе («петушиный шаг»). Эти симптомы являются частью синдромамозжечковой атаксии.
Вестибулярные рефлексы, их характеристика. Нистагм глаз.
Статические и статокинетические рефлексы. Равновесие поддерживается рефлекторно, без принципиального участия в этом сознания. Выделяют статические и статокинетические рефлексы. Вестибулярные рецепторы и соматосенсорные афференты, особенно от проприоцепторов шейной области, связаны и с теми и с другими. Статические рефлексы обеспечивают адекватное взаиморасположение конечностей, а также устойчивую ориентацию тела в пространстве, т.е. позные рефлексы. Вестибулярная афферентация поступает в данном случае от отолитовых органов. Статический рефлекс, легко наблюдаемый у кошки благодаря вертикальной форме ее зрачка, – компенсаторное вращение глазного яблока при повороте головы вокруг длинной оси тела (например, левым ухом вниз). Зрачки при этом все время сохраняют положение, очень близкое к вертикальному. Такой рефлекс наблюдается и у человека. Статокинетические рефлексы – это реакции на двигательные стимулы, сами выражающиеся в движениях. Они вызываются возбуждением рецепторов полукружных каналов и отолитовых органов; их примеры – вращение тела кошки в падении, обеспечивающее ее приземление на все четыре лапы, или движения человека, восстанавливающего равновесие после того, как он споткнулся.
Один из статокинетических рефлексов – вестибулярный нистагм. Как говорилось выше, вестибулярная система вызывает различные движения глаз; нистагм как их особая форма наблюдается в начале более интенсивного, чем обычные короткие повороты головы, вращения. При этом глаза поворачиваются против направления вращения, чтобы удержать исходное изображение на сетчатке, однако, не достигая своего крайнего возможного положения, резко «перескакивают» в направлении вращения, и в поле зрения оказывается другой участок пространства. Затем следует их медленное возвратное движение.
Медленная фаза нистагма запускается вестибулярной системой, а быстрый «перескок» взгляда–предмостовой частью ретикулярной формации.
При вращении тела вокруг вертикальной оси раздражаются практически только горизонтальные полукружные каналы, т. е. отклонение их купул вызывает горизонтальный нистагм. Направление обоих его компонентов (быстрого и медленного) зависит от направления вращения и, таким образом, от направления деформации купул. Если тело вращается вокруг горизонтальной оси (например, проходящей через уши или саггитально через лоб), стимулируются вертикальные полукружные каналы и возникает вертикальный, или вращательный, нистагм. Направление нистагма принято определять по его быстрой фазе, т.е. при «правом нистагме» взгляд «перескакивает» вправо.
При пассивном вращении тела к возникновению нистагма ведут два фактора: стимуляция вестибулярного аппарата и перемещение поля зрения относительно человека. Оптокинетический (вызванный зрительной афферентацией) и вестибулярный нистагмы действуют синергически.
Диагностическое значение нистагма. Нистагм используется в клинике для тестирования вестибулярной функции. Испытуемый сидит в специальном кресле, которое длительное время вращается с постоянной скоростью, а затем резко останавливается. Остановка вызывает отклонение купулы в направлении, противоположном тому, в котором она отклонялась в начале движения; результат–нистагм. Его направление можно определить, регистрируя деформацию купулы; оно должно быть противоположным направлению предшествующего движения. Запись движений глаз напоминает получаемую в случае оптокинетического нистагма. Она называется нистагмограммой.
Проведя тест на поствращательный нистагм, важно устранить возможность фиксации взгляда в одной точке, поскольку при глазодвигательных реакциях зрительная афферентация доминирует над вестибулярной и в некоторых условиях способна подавить нистагм. Поэтому испытуемому надевают очки Френцеля с сильновыпуклыми линзами и встроенным источником света. Они делают его «близоруким» и неспособным фиксировать взор, одновременно позволяя врачу без труда наблюдать движения глаз. Такие очки необходимы и в тесте на наличие спонтанного нистагма–первой, простейшей и наиболее важной процедуре при клиническом исследовании вестибулярной функции.
Еще один клинический способ запуска вестибулярного нистагма – термостимуляция горизонтальных полукружных каналов. Его преимущество–в возможности тестировать каждую сторону тела отдельно. Голову сидящего испытуемого отклоняют назад приблизительно на 60° (у лежащего на спине человека ее приподнимают на 30°), чтобы горизонтальный полукружный канал занимал строго вертикальное направление. Затем наружный слуховой проход промывают холодной или теплой водой. Наружный край полукружного канала расположен к нему очень близко, поэтому сразу же охлаждается или нагревается. В соответствии с теорией Барани плотность эндолимфы при нагревании понижается; следовательно, ее нагретая часть поднимается, создавая разность давлений по обе стороны купулы; возникающая деформация вызывает нистагм. Исходя из его природы, этот вид нистагма называют калорическим. При нагревании он направлен к месту термического воздействия, при охлаждении–в обратную сторону. У людей, страдающих вестибулярными расстройствами, нистагм отличается от нормального качественно и количественно. Детали его тестирования приведены в работе. Следует отметить, что калорический нистагм может возникать в космических кораблях в условиях невесомости, когда различия плотности эндолимфы несущественны. Следовательно, в его запуске участвует, по крайней мере, еще один, пока не известный механизм, например, прямое термическое воздействие на вестибулярный орган.
Функцию отолитового аппарата можно тестировать, наблюдая глазодвигательные реакции при наклонах головы или при возвратно–поступательных движениях пациента, находящегося на специальной платформе.
Строение и функции наружного, среднего и внутреннего уха. Механизм слуховой рецепции.
Строение и функции структур наружного и среднего уха.
Наружнее ухо состоит из:
Ушной раковины
Наружного слухового прохода
Внешней стороны барабанной перепонки
Функции наружного уха:
Направленный прием звуковых волн
Концентрация звука в направлении наружного слухового прохода
Ограничение потока звуковых сигналов, поступающих с тыла головы
Защитная функция (сера)
Согревание и увлажнение воздуха
Среднее ухо состоит из:
Барабанная перепонка - уменьшает амплитуду колебаний
Молоточек, наковальня, стремечко – увеличивают силу
Овальное окно (ведет во внутреннее ухо)
Круглое окно (чтобы колебания не затухали, т.к. жидкость по своим свойствам – несжимаема)
Евстахиева труба (соединение среднего уха с носоглоткой, выравнивает давление)
!!! Поверхность стремечка в 22 раз меньше БП, во столько же раз увеличивается давление стремечка на овальное окно, при этом даже слабые звуковые волны могут вызвать колебания жидкости в улитке
Мышцы:
Мышца, напрягающая барабанную перепонку (m. tensor tympani)
Усиливает напряжение БП при сокращении
Ограничивает амплитуды колебаний при сильном звуке
Стременная мышца (m.stapedius)
Фиксирует стремечко и ограничивает его движения
Сокращаются при – глотании, жевании, зевании, речевой деятельности, т.е. участвуют в ориентации и в реакции обратной связи от речевой системы к слуховому входу.
(+) Рефлекторное сокращение этих мышц наступает через 10 сек после сильного звука и зависит от его амплитуды (автоматическое предохранение от перегрузки)
Строение и функции структур внутреннего уха.
Внутреннее ухо – сообщается с полостями и каналами (где находится запас жидкости) в лабиринте височной кости.
Улитка – костный спиральный канал (2.5 витка), разделен;
Преддверной (вестибулярной) мембраной – мембрана Рейснера
Основной (базиллярной) мембраной
Эти мембраны делят улитку на 3 части (хода):
Верхний канал (лестница преддверия- scala vestibuli)
Сообщается с нижним каналом улитки через овальное отверстие
Заполнен перилимфой
Нижний канал (барабанная лестница – scala tympany) – схож с верхним
Средний канал (перепончатый канал – scala media)
Не сообщается с другими каналами
Заполнен эндолимфой, имеющий положительный заряд(в 100 раз больше К)
Место расположения кортиева органа, который содержит рецепторы – волосковые клетки (вторично-чувствующие механо-Re)
Вторично-чувствующие механо-Re – преобразуют механические колебания в БЭП. Состоят из 2 слоев:
Внутренний – 1 ряд (3500 шт)
Наружный – 3-5 рядов (12000-25000 шт)
Каждая рецепторная клетка имеет сотни волосков.
Волоски прикрыты покровной мембраной (из соединительной ткани), который закрывает один конец, а второй свободен, и они скользят по клеткам рецептора.
Движение звуковой волны:
Колебание барабанной перепонки
Колебание и усиление волны по костному молоточку
Наковальня-> стремечко -> овальное окно
Колебание перилимфы в верхнем улитковом канале
Распространение колебаний по эндолимфе (бегущая волна)
Передача на перилимфу нижнего канала
Круглое окно
Механизм возбуждения слуховых рецепторов
Поступление колебательных звуковых движений в слуховой проход
Колебания барабанной перепонки
Колебания костного аппарата
Колебания овального отверстия
Возникновение колебания в полостях вестибулярной и вестибулярной лестниц
Колебание эндолимфы в среднем канале
Колебание тинкториальной мембраны, переход на базальную
Колебания волосковой клетки и тинкториальной мембраны
Волоски склоняются и возникает возбуждение
Открываются механо-управляемые К-каналы (К в эгдолимфе больше в 100 раз)
Калий идет в волосковую клетку по градиенту концентрации
Возникает рецепторный потенциал
Открываются потенциал-управляемые Са-каналы
Вход Са в клетку
Выход медиаторов – глутамат (аспартат)
Генерация ВПСП
Типы электрических потенциалов:
Микрофонный – возникает в результате деформации волосковых клеток кортиева органа, полностью воспроизводит форму и частоту звуковой волны ( до 4-5 кГц).
Рецепторный потенциал – электрическое возбуждение рецепторных клеток
Суммационный потенциал (свыше 4-5 кГц) – сдвиг постоянного потенциала улитки, слабо выраженный при низких частотах.
Величина(пол./отриц.) – пропорциональна интенсивности звука
Рецепторный потенциал – возбуждение в окончаниях слуховых нейронов
Эндокохлеарный потенциал – постоянный потенциал улитки. Источник – активный секреторный процесс сосудистой полости клеток эндолимфы.
Звук -> микрофонный и суммационный потенциал -> генеративный потенциал
Проводниковый и корковый отделы слуховой сенсорной системы. Бинауральный слух.
Проводниковый и корковый отделы слухового анализатора.
I нейрон – дендриты от волосковых клеток
Спинальный ганглий, образуют нервус вестибулокохлеарис (улитковую часть)
II нейрон – переключение на кохлеарные ядра в продолговатом мозге
III нейрон – верхние оливарные ядра (I перекрест), трапецевидное тело ядра
IV нейрон – в составе латеральной петли в нижние бугры 4-холмия (II перекрест)
V нейрон – специфические ядра таламуса (медиальное коленчатое тело)
Слуховая зона коры верхней части височной доли (I проекционное поле)
Ассоциативная область коры большого мозга (II проекционное поле)
1)Нервные импульсы от коленчатого тела в основном идут в первичную слуховую кору.
2)Вторичная слуховая кора – активируется таламусом и первичной слуховой корой
3)При взаимодействии первичной, вторичной коры и ассоциативной коры (третичная кора) – формируется ощущение.
Бинауральный слух.
Человек и животные обладают пространственным слухом, т.е. способностью определять положение источника звука в пространстве. Это свойство основано на наличии бинаурального слуха, или слушания двумя ушами. Для него важно и наличие двух симметричных половин на всех уровнях слуховой системы. Острота бинаурального слуха у человека очень высока: положение источника звука определяется с точностью до 1 углового градуса. Основой этого служит способность нейронов слуховой системы оценивать интерауральные (межушные) различия времени прихода звука на правое и левое ухо и интенсивности звука на каждом ухе. Если источник звука находится в стороне от средней линии головы, звуковая волна приходит на одно ухо несколько раньше и имеет большую силу, чем на другом ухе. Оценка удаленности источника звука от организма связана с ослаблением звука и изменением его тембра.
При раздельной стимуляции правого и левого уха через наушники задержка между звуками уже в 11 мкс или различие в интенсивности двух звуков на 1 дБ приводят к кажущемуся сдвигу локализации источника звука от средней линии в сторону более раннего или более сильного звука. В слуховых центрах есть нейроны с острой настройкой на определенный диапазон интерауральных различий по времени и интенсивности. Найдены также клетки, реагирующие лишь на определенное направление движения источника звука в пространстве.
Методы исследования слуха. Исследование костной и воздушной проводимости (пробы Вебера и Ринне).
№ |
Название метода |
Характеристика |
Объективные методы |
||
1 |
Безусловно-рефлекторные |
предназначены для обследования слуха, как правило, детей в возрасте до 5 лет, так как в этом случае общепринятые аудиометрические методы не могут быть применены. Методы этой группы основаны на проявлении безусловных ориентировочных рефлексов в ответ на звуковое раздражение. Это такие рефлексы, как общие мышечные сокращения, мимические реакции, реакции глаз (мигание, зажмуривание и раскрытие, расширение зрачка), движения головы, туловища, конечностей, сосательные рефлексы грудных детей, сердечно-сосудистые и дыхательные рефлексы. Указанные методы применяются при интенсивности звукового раздражителя 80-90 дБ, что позволяет выявлять в основном большие и средние потери слуха. Вместе с тем у большинства детей наблюдается расширение зрачков даже при незначительном превышении пороговой слышимости. При использовании безусловно-рефлекторных методов следует иметь в виду наличие значительного индивидуального разброса и непостоянство реакций ребенка. |
2 |
Условно-рефлекторные |
также применяются для обследования слуха у маленьких детей. В их основе - проявление рефлекса на какое-либо действие, совершаемое одновременно с подачей звука. В результате с большой достоверностью возникает устойчивая условная реакция на звук даже при небольшом превышении порога восприятия. Например, если сочетать в течение 2-3 недель кормление грудного ребенка с подачей звука, то наблюдается сосательный рефлекс в ответ только на звуковое раздражение. Отсутствие рефлекса при подаче звука свидетельствует о потере слуха. Для выработки условного рефлекса на мигание глаз при действии звуков можно использовать слабую струю теплого воздуха, направляемую в лицо одновременно со звуком. |
3 |
Анализ слуховых вызванных потенциалов |
одна из наиболее применяемых в настоящее время методик объективного исследования слуха. В ответ на звуковой сигнал в различных отделах слухового анализатора - от улитки до коры головного мозга - возникают электрические сигналы, составляющие миллионные доли вольта. Чтобы их зарегистрировать, используют высокочувствительные усилители, а также многократное повторение звуковых сигналов. Таким образом, с помощью компьютера производится накопление, суммирование и усреднение регистрируемых сигналов слуховой системы. Электроды, регистрирующие потенциалы, располагаются на макушке головы и сосцевидных отростках. Все вызванные потенциалы в зависимости от латентного периода (времени от начала стимуляции до начала ответной реакции) делятся на 3 группы: коротколатентные сигналы (до 12 мсек) отражают возбуждение улитки и слухового нерва, среднелатентные (12-50 мсек) и длиннолатентные (50-300 мсек) отражают ответ различных участков коры головного мозга. Каждый пик потенциалов указывает на конкретные части слуховой системы или головного мозга. |
Субъективные методы |
||
1 |
Обследование с помощью шепотной и разговорной речи |
Это самый простой метод. При обследовании необходимо предъявлять известные ребенку слова. Если ребенок владеет только лепетными словами («мяу» - кошка, «туту» - автомобиль и т. д.), то надо их использовать. Если ребенок не понимает слов, то выявляется способность воспринимать гласные и согласные, такие как низкочастотные о, у, л, м, н, р, б, т, к, в, п или высокочастотные - и, а, е, с, г, х, шипящие. При этом большинство гласных и высокочастотные согласные слышатся лучше и воспринимаются на более далеких расстояниях, чем низкочастотные согласные. Обследование необходимо проводить эмоционально, выражая удовлетворение или удивление при правильном восприятии слова. Активизирует занятия показ картинок, соответствующих словам, использование игровых моментов. Слова произносятся после спокойного вдоха, на резервном воздухе легких, в этом случае легко уравнять громкость и скорость их произнесения у разных людей. Речевой материал необходимо предъявлять непосредственно у уха и затем увеличивать расстояние. Шепотная речь при нормальном слухе воспринимается с 5-10 м для низкочастотных звуков и с 15-20 м - для высокочастотных. Для детей старше 5 лет целесообразно использовать специальные таблицы, соответствующие возрасту, а также исследовать «фонематический слух», т. е. возможность различать слова, отличающиеся только одной буквой (фонемой) (Миша - Маша, бочка - почка, дочка - точка). Изучение слуха с помощью шепотной и разговорной речи дает оценочное представление о слухе, более детально слуховая функция анализируется последующими методами. |
2 |
Пороговая тональная аудиометрия |
В настоящее время это один из основных методов исследования слуха, при котором используется специальная электроакустическая аппаратура - аудиометры. Пороговая аудиометрия заключается в определении на разных частотах наименьшей интенсивности звукового сигнала, при которой возникает ощущение звука. Обследование необходимо проводить в звукоизолированном помещении, интенсивность окружающего шума не должна превышать 20 дБ. На каждой частоте вначале подается громкий сигнал, чтобы испытуемый имел представление, какой звук он должен услышать. Затем сила звука уменьшается до неслышимого, после чего плавно увеличивается до появления ощущения звука. Далее интенсивность звука уменьшается на 10 дБ и вновь дискретно повышается с остановкой на 10 с через каждые 5 дБ до возникновения ощущения звука. Значения пороговой интенсивности тона для каждой из исследуемых частот откладываются на аудиограмме. Время аудиометрического oбследования не должно превышать 60 мин во избежание утомления и ослабления внимания. |
3 |
Пороговая тональная аудиометрия по воздушному каналу |
Звук подается на ухо через наушники. Исследование начинается с тона 1000 Гц, который является резонансным тоном проводящей системы уха и поэтому комфортен для слуха. После этой частоты переходят к определению порогов по низким частотам - 500, 250, 125 Гц, как лучше сохранившимся, а затем по высоким - 2000, 4000, 8000 Гц. Исследование заканчивается на частоте 1000 Гц, чтобы подтвердить начальный результат. Результаты отмечаются на аудиограмме: по правому уху кружком (о) или красным цветом, по левому - крестиком (х), синим или зеленым цветом. Аудиограммы воздушного звукопроведения обозначают сплошной линией, костного звукопроведения - пунктиром. |
4 |
Речевая аудиометрия |
Применяется при обследовании людей, имеющих развитую речь. Цель речевой аудиометрии - определение величины потери слуха на речь. Записанный на магнитофон набор слов подается испытуемому через аудиометр. После каждого слова следует пауза, в течение которой он должен ответить, что услышал. Экспериментатор считает число правильно воспроизведенных слов. Если их число оказывается недостаточным, громкость прибавляется на 5 дБ и обследование повторяется. |
Исследование костной и воздушной проводимости
1. Опыт Ринне заключается в сравнении воздушной и костной проводимости. Звучащий камертон С128 ставят на сосцевидный отросток исследуемого и, включив секундомер, замечают, сколько времени он звучал. По прекращении звучания на сосцевидном отростке подносят камертон к отверстию слухового прохода. У здорового человека проводимость через воздух больше проводимости через кость - это обозначают как «положительный опыт Ринне». При наличии же поражения в среднем ухе или вообще звукопроводящего аппарата опыт Ринне может быть отрицательным, т. е. звучание с кости будет продолжительнее звучания через воздух; обычно это указывает на заболевание звукопроводящего аппарата.
2. Опыт Вебера производится так. Звучащий камертон помещают на темя больного и спрашивают его, в каком ухе он слышит звучание. При здоровом состоянии ушей исследуемый слышит звучание в голове, не относя звук ни к одному из ушей. При нарушении звукопроводящего аппарата звук слышится в больном ухе, при нарушении звуковоспринимающего аппарата он слышен в здоровом ухе. Известно несколько попыток дать объяснение усилению костной проводимости при заболевании среднего уха. Некоторые указывают, что при здоровом состоянии ушей звуковые волны от звучащего камертона, беспрепятственно распространяясь по черепу, как бы выходят через уши в окружающую среду и не задерживаются в каком-либо ухе. При наличии препятствия в виде воспалительного процесса среднего уха или инородного тела (серная пробка) в слуховом проходе звуковые волны, отражаясь от препятствия, как бы снова ударяют в звуковоспринимающий аппарат внутреннего уха и звучат в больном ухе. При поражении же звуковоспринимающего аппарата звук может появиться только в здоровом ухе. Так, Бецольд считает, что при заболеваниях звукопроводящего аппарата ограничение движений слуховых косточек создает условия для худшей передачи через воздух, чем через кость.
Л. Е. Комендантов и другие авторы объясняли укорочение восприятия звучания камертона через кость на здоровой стороне утомлением нерва.
Г. Г. Куликовский, исследуя слуховую функцию больных в звуконепроницаемой камере, зарегистрировал незначительное укорочение костной проводимости при поражении звукопроводящего аппарата. Он считает, что наблюдающееся в обычных условиях исследования слуха удлинение костной проводимости у этого рода больных зависит от неблагоприятных в акустическом отношении условий восприятия звука.
При поражении мозга и его оболочек латеризации звука в опыте Вебера не наблюдается, если при этом нет нарушения слуховой функции.
Состав и функции оптического аппарата глаза. Аккомодация глаза, ее механизмы при рассматривании близких и далеких предметов.
Зрение эволюционно приспособлено к восприятию электромагнитных излучений в определенной, весьма узкой части их диапазона (видимый свет). Зрительная система дает мозгу более 90% сенсорной информации. Зрение — многозвеньевой процесс, начинающийся с проекции изображения на сетчатку уникального периферического оптического прибора — глаза. Затем происходят возбуждение фоторецепторов, передача и преобразование зрительной информации в нейронных слоях зрительной системы, а заканчивается зрительное восприятие принятием высшими корковыми отделами этой системы решения о зрительном образе.
Состав и функции оптического аппарата глаза.
Глазное яблоко имеет шарообразную форму, что облегчает его повороты для наведения на рассматриваемый объект. На пути к светочувствительной оболочке глаза (сетчатке) лучи света проходят через несколько прозрачных сред — роговицу, хрусталик и стекловидное тело. Определенная кривизна и показатель преломления роговицы и в меньшей мере хрусталика определяют преломление световых лучей внутри глаза.
Преломляющую силу любой оптической системы выражают в диоптриях (D). Одна диоптрия равна преломляющей силе линзы с фокусным расстоянием 100 см. Преломляющая сила здорового глаза составляет 59D при рассматривании далеких и 70.5D — при рассматривании близких предметов. Чтобы схематически представить проекцию изображения предмета на сетчатку, нужно провести линии от его концов через узловую точку (в 7 мм сзади от роговой оболочки). На сетчатке получается изображение, резко уменьшенное и перевернутое вверх ногами и справа налево
Аккомодация, ее механизм.
Аккомодацией называют приспособление глаза к ясному видению объектов, удаленных на разное расстояние. Для ясного видения объекта необходимо, чтобы он был сфокусирован на сетчатке, т. е. чтобы лучи от всех точек его поверхности проецировались на поверхность сетчатки (рис. 14.4). Когда мы смотрим на далекие предметы (А), их изображение (а) сфокусировано на сетчатке и они видны ясно. Зато изображение (б) близких предметов (Б) при этом расплывчато, так как лучи от них собираются за сетчаткой. Главную роль в аккомодации играет хрусталик, изменяющий свою кривизну и, следовательно, преломляющую способность. При рассматривании близких предметов хрусталик делается более выпуклым (см. рис. 14.2), благодаря чему лучи, расходящиеся от какой-либо точки объекта, сходятся на сетчатке. Механизмом аккомодации является сокращение ресничных мышц, которые изменяют выпуклость хрусталика. Хрусталик заключен в тонкую прозрачную капсулу, которую всегда растягивают, т. е. уплощают, волокна ресничного пояска (циннова связка). Сокращение гладких мышечных клеток ресничного тела уменьшает тягу цинновых связок, что увеличивает выпуклость хрусталика в силу его эластичности. Ресничные мышцы иннервируются парасимпатическими волокнами глазодвигательного нерва. Введение в глаз атропина вызывает нарушение передачи возбуждения к этой мышце, ограничивает аккомодацию глаза при рассматривании близких предметов. Наоборот, парасимпатомиметические вещества — пилокарпин и эзерин — вызывают сокращение этой мышцы.
Близорукость, дальнозоркость, астигматизм.
Их происхождение и способы коррекции.
Близорукость, ее происхождение и способ коррекции.
Миопия или близорукость — параллельные лучи фокусируются не на сетчатке, а впереди нее. Это возникает при чрезмерно большой длине глазного яблока или преломляющей силе глаза. Близкие предметы видятся хорошо, удаленные — расплывчато. Коррекция: использование рассеивающих двояковогнутых линз.

Дальнозоркость, ее происхождение и способ коррекции.
Гиперметропия или дальнозоркость — параллельные лучи фокусируются за сетчаткой. Это возникает из-за малой длины глазного яблока или слабой преломляющей способности глаза. Коррекция: использование собирающих двояковыпуклых линз.

Астигматизм, ее происхождение и способ коррекции.
Неодинаковое преломление лучей в разных направлениях (например, по горизонтальному и вертикальному меридиану). Следует отнести к несовершенству строения глаза как оптического инструмента.
Астигматизм обусловлен тем, что роговая оболочка не является строго сферической поверхностью: в различных направлениях она имеет различный радиус кривизны. При сильных степенях астигматизма эта поверхность приближается к цилиндрической, что дает искаженное изображение на сетчатке. Коррекция: специальные цилиндрические стекла (если, например, роговая оболочка преломляет слабее в вертикальном направлении, то стекло должно преломлять в этом направлении сильнее).
Зрачковый рефлекс, механизмы сужения и расширения зрачка.
Зрачком называют отверстие в центре радужной оболочки, через которое лучи света проходят внутрь глаза. Зрачок повышает четкость изображения на сетчатке, увеличивая глубину резкости глаза. Пропуская только центральные лучи, он улучшает изображение на сетчатке также за счет устранения сферической аберрации. Если прикрыть глаз от света, а затем открыть его, то расширившийся при затемнении зрачок быстро сужается («зрачковый рефлекс»). Мышцы радужной оболочки изменяют величину зрачка, регулируя поток света, попадающий в глаз. Так, на очень ярком свету зрачок имеет минимальный диаметр (1,8 мм), при средней дневной освещенности он расширяется (2,4 мм), а в темноте расширение максимально (7,5 мм). Это приводит к ухудшению качества изображения на сетчатке, но увеличивает чувствительность зрения. Предельное изменение диаметра зрачка изменяет его площадь примерно в 17 раз. Во столько же раз меняется при этом световой поток. Между интенсивностью освещения и диаметром зрачка имеется логарифмическая зависимость. Реакция зрачка на изменение освещенности имеет адаптивный характер, так как в небольшом диапазоне стабилизирует освещенность сетчатки.
В радужной оболочке имеется два вида мышечных волокон, окружающих зрачок: кольцевые (m. sphincter iridis), иннервируемые парасимпатическими волокнами глазодвигательного нерва, а также радиальные (m. dilatator iridis), иннервируемые симпатическими нервами. Сокращение первых вызывает сужение, сокращение вторых — расширение зрачка. Соответственно этому ацетилхолин и эзерин вызывают сужение, а адреналин — расширение зрачка. Зрачки расширяются во время боли, при гипоксии, а также при эмоциях, усиливающих возбуждение симпатической системы (страх, ярость). Расширение зрачков — важный симптом ряда патологических состояний, например болевого шока, гипоксии.
У здоровых людей размеры зрачков обоих глаз одинаковые. При освещении одного глаза зрачок другого тоже суживается; такая реакция называется содружественной. В некоторых патологических случаях размеры зрачков обоих глаз различны (анизокория).
Строение и функции сетчатки глаза. Пигментный слой сетчатки глаза, его функции.
Сетчатка – периферический рецептор зрительного анализатора, специализированная часть мозговой коры, вынесенная на периферию. Выстилает всю внутреннюю поверхность сосудистого тракта, состоит из двух отделов:
1) оптическая часть – задние 2/3 сетчатки, высокодифференцированная нервная ткань из 10-и слоев, заканчивающаяся у места перехода цилиарного тела в хориоидею
2) слепая часть – передняя 1/3 сетчатки, малодифференцированная нервная ткань из 2-х слоев, продолжается до зрачкового края, где образует краевую пигментную кайму.
Место выхода зрительного нерва – диск зрительного нерва, на расстоянии около четырех мм от диска зрительного нерва имеется углубление – желтое пятно.
Гистологически сетчатка – цепь трех нейронов: наружного – фоторецепторного, среднего – ассоциативного, внутреннего – ганглионарного, образующих 10 слоев сетчатки:
1) слой пигментного эпителия – клетки в виде шестигранных призм, расположенных в один ряд, тела клеток заполнены зернами пигмента – фусцина; плотно спаян с сетчаткой
2) слой палочек и колбочек – светочувствительный слой, наружные сегменты фоторецепторов (палочек и колбочек). Палочки – тонкие, цилиндрические, содержат пигмент родопсин, являются аппаратом сумеречного зрения, их количество в 20 раз выше количества колбочек. Колбочки – конусообразные, толще палочек, содержат пигмент йодопсин, являются аппаратом центрального и цветового зрения. В области желтого пятна находятся только колбочки.
3) наружная глиальная пограничная мембрана – полоса межклеточный сцеплений
4) наружный зернистый (ядерный) слой – образован ядрами фоторецепторов
5) наружный сетчатый слой – содержит синапсы, обеспечивающие связь первого и второго нейронов
6) внутренний зернистый (ядерный) слой – тела и ядра вторых биполярных нейронов, имеющих два отростка – один для связи с фотосенсорными клетками, второй – для образования синапса с дендритами оптико-ганглионарных клеток. Биполяры контактируют с несколькими палочковыми клетками и только с одной из колбочковых клеток.
7) внутренний сетчатый слой – синапсы биполярных и оптико-ганглионарных нейронов.
8) ганглионарный слой – оптико-ганглионарные нейроны, имеют крупное ядро, сильно ветвящиеся дендриты и один аксон – цилиндр.
9) слой нервных волокон – аксоны оптико-ганглионарных нейронов, формирующих зрительный нерв
10) внутренняя глиальная пограничная мембрана – покрывает поверхность сетчатки изнутри, основная мембрана, основаниями отросткой нейроглиальных клеток Мюллера.
Пигментный слой сетчатки глаза. Слой пигментных эпителиоцитов сетчатой оболочки глаза включает около 6 миллионов пигментных клеток, которые своей базальной поверхностью лежат на базальной мембране сосудистой оболочки. Светлая цитоплазма пигментных клеток (меланоцитов) бедна органеллами общего значения, содержит большое количество пигмента (меланосом). Ядра меланоцитов имеют сферическую форму. От апикальной поверхности меланоцитов отходят отростки (микроворсинки), которые заходят между концами палочек и колбочек. Каждую палочку окружают 6-7 таких отростков, каждую колбочку — 40 отростков. Пигмент этих клеток способен мигрировать из тела клетки в отростки, а из отростков в тело меланоцита. Эта миграция осуществляется под влиянием меланоцитостимулируюгцего гормона промежуточной части аденогипофиза и при участии филаментов внутри самой клетки.
Функции пигментного слоя сетчатки многочисленны:
1) является составной частью адаптационного аппарата глаза;
2) участвует в торможении перекисного окисления;
3) выполняет фагоцитарную функцию;
4) участвует в обмене витамина А.
Фоторецепторы, их классификация, строение и функции.
К пигментному слою изнутри примыкает слой фоторецепторов: палочек и колбочек. В сетчатке каждого глаза человека находится 6—7 млн колбочек и 110—123 млн палочек. Они распределены в сетчатке неравномерно. Центральная ямка сетчатки (fovea centralis) содержит только колбочки (до 140 тыс. на 1 мм2). По направлению к периферии сетчатки их число уменьшается, а число палочек возрастает, так что на дальней периферии имеются только палочки. Колбочки функционируют в условиях больших освещенностей, они обеспечивают дневное . и цветовое зрение; намного более светочувствительные палочки ответственны за сумеречное зрение.
Цвет воспринимается лучше всего при действии света на центральную ямку сетчатки, где расположены почти исключительно колбочки. Здесь же и наибольшая острота зрения. По мере удаления от центра сетчатки восприятие цвета и пространственное разрешение становятся все хуже. Периферия сетчатки, где находятся исключительно палочки, не воспринимает цвета. Зато световая чувствительность колбочкового аппарата сетчатки во много раз меньше, чем палочкового, поэтому в сумерках из-за резкого понижения «колбочкового» зрения и преобладания «периферического» зрения мы не различаем цвет («ночью все кошки серы»).
Нарушение функции палочек, возникающее при недостатке в пище витамина А, вызывает расстройство сумеречного зрения — так называемую куриную слепоту: человек совершенно слепнет в сумерках, но днем зрение остается нормальным. Наоборот, при поражении" колбочек возникает светобоязнь: человек видит при слабом" свете, но слепнет при ярком освещении. В этом случае может развиться и полная цветовая слепота — ахромазия.
Строение фоторецепторной клетки.
К слою пигментного эпителия изнутри примыкает слой фоторецепторов, которые своими светочувствительными члениками обращены в сторону, противоположную свету.
Каждый фоторецептор - палочка или колбочка - состоит из чувствительного к действию света наружного сегмента, содержащего зрительный пигмент, и внутреннего сегмента, содержащего ядро и митохондрии, обеспечивающие энергетические процессы в фоторецепторной клетке.
Электронно-микроскопические исследования выявили, что наружный сегмент каждой палочки состоит из 400800 тонких пластинок, или дисков, диаметром около 6 мкм (6-10--6 м). Каждый диск представляет собой двойную мембрану, состоящую из мономолекулярных слоев липидов, находящихся между слоями молекул белка. С молекулами белка связан ретиналь, входящий в состав зрительного пигмента родопсина.
Наружный и внутренний сегменты фоторецепторной клетки разделены мембранами, через которые проходит пучок из 16-18 тонких фибрилл. Внутренний сегмент переходит в отросток, с помощью которого фоторецепторная клетка передает возбуждение через синапс на контактирующую с ней биполярную нервную клетку.
Зрительные пигменты, их виды и функции. Фотохимические процессы в рецепторах сетчатки глаза.
- Родопсин, или зрительный пурпур: содержится в палочках и представляет собой высокомолекулярное соединение (молекулярная масса 270 000), состоящее из ретиналя - альдегида витамина А и белка опсина. При действии кванта света происходит цикл фотофизических и фотохимических превращений этого вещества: ретиналь изомеризуется, его боковая цепь выпрямляется, связь ретиналя с белком нарушается, активируются ферментативные центры белковой молекулы. После чего ретиналь отщепляется от опсина. Под влиянием фермента, названного редуктазой ретиналя, последний переходит в витамин А. При затемнении глаз происходит регенерация зрительного пурпура, т.е. ресинтез родопсина. Для этого процесса необходимо, чтобы сетчатка получала цис-изомер витамина А, из которого образуется ретиналь. Если же витамин А в организме отсутствует, образование родопсина резко нарушается, что и приводит к развитию упомянутой выше куриной слепоты.
- Йодопсин: содержится в колбочках и представляет собой также соединение ретиналя с белком опсином, который образуется в колбочках и отличается от опсина палочек в наибольшей степени поглощает желтый свет с длиной волны около 560 нм. В колбочках имеются также пигменты хлоролаб (лучи зеленой части спектра), эритролаб (лучи красной части спектра) и цианолаб (лучи синей части спектра).
Фотохимические процессы в рецепторах сетчатки глаза.
При действии кванта света в рецепторах сетчатки происходит цепь фотохимических реакций, связанных с распадом зрительных пигментов родопсина и йодопсина и их ресинтез в темноте. Родопсин — пигмент палочек, высокомолекулярное соединение, состоящее из ретиналя — альдегида витамина А и белка опсина. При поглощении кванта света молекулой родопсина 11-цис-ретиналь выпрямляется и превращается в транс-ретиналь; белковая часть молекулы обесцвечивается и переходит в состояние метародопсина II, который взаимодействует с примембранным белком гуанозинтрифосфат-связанным белком трансдуцином. Последний запускает реакцию обмена ГДФ на ГТФ, что приводит к усилению светового сигнала. ГТФ вместе с трансдуцином активирует молекулу примембранного белка — фермента фосфодиэстеразы (ФДЭ), который разрушает молекулу цГМФ, вызывая еще большее усиление светового сигнала. Падает содержание цГМФ и закрываются каналы для Na+ и Са+ —> возникает гиперполяризация мембраны фоторецептора —> возникает рецепторный потенциал.
Морфофункциональная характеристика проводникового и коркового отделов зрительной сенсорной системы. Бинокулярное зрение, его происхождение.
Проводниковый отдел зрительного анализатора.
Проводниковый отдел зрительного анализатора состоит из волокон зрительного нерва, соединяющих сетчатку с высшими зрительными центрами.
Зрительные пути. В настоящее время большинство ученых придерживается точки зрения, что зрительный путь состоит из 4 нейронов. Первый нейрон - палочки и колбочки, второй - биполярные клетки, третий - мультиполярные клетки сетчатки и их аксоны. Мультиполярные клетки наружного коленчатого тела дают начало 4 нейрону зрительного пути. Зрительный путь (рис.7) соединяет сетчатку с головным мозгом. Различают пять частей зрительного пути:
1. зрительный нерв;
2. зрительный перекрест;
3. зрительный тракт;
4. латеральное коленчатое тело;
5. зрительный центр восприятия.
Зрительный нерв относится к черепным нервам (II пара). Нерв образован из аксонов мультиполярных клеток, которые доходят до наружного коленчатого тела и центробежных волокон, являющихся элементами обратной связи. В составе зрительного нерва имеется около 1млн. волокон, что примерно соответствует количеству рецепторных полей сетчатки (около 800 тыс.). В области турецкого седла зрительные нервы сливаются друг с другом и образуют зрительный перекрест.
Зрительный перекрест. В зрительном перекресте совершаются расслоение и частичный перекрест волокон зрительного нерва. Перекрещиваются волокна, идущие от внутренних половин сетчатки. Волокна, идущие от височных половин сетчатки, располагаются по наружным сторонам перекреста. Волокна, исходящие из желтого пятна перекрещиваются лишь частично. От зрительного перекреста начинаются зрительные тракты. После частичного перекреста зрительных нервов образуется правый и левый зрительные тракты.
Зрительный тракт. Начинаясь у задней поверхности зрительного перекреста, зрительный тракт заканчивается в основном (80%) в латеральном коленчатом теле промежуточного мозга, но часть волокон доходит до подушки таламуса и до верхнего холмика четверохолмия крыши среднего мозга.
В латеральном коленчатом теле заканчивается периферический нейрон и берет начало центральный нейрон зрительного пути, который после выхода из латерального коленчатого тела в виде зрительной лучистости (пучок Грациоле) направляется в кортикальные зрительные центры.
Корковый отдел зрительного анализатора.
Центральный отдел зрительного анализатора
Центральный отдел зрительного анализатора условно можно разделить на 2 части:
1 - ядро зрительного анализатора первой сигнальной системы - в области шпорной борозды, что в основном соответствует полю 17 коры головного мозга по Бродману);
2 - ядро зрительного анализатора второй сигнальной системы - в области левой угловой извилины.
Поле 17 в основном созревает к 3 - 4 годам. Оно является органом высшего синтеза и анализа световых раздражителей. При поражении поля 17 может наступить физиологическая слепота. К центральному отделу зрительного анализатора относятся поля 18 и 19, где обнаружены зоны с полным представительством поля зрения. Кроме того, нейроны, реагирующие на зрительную стимуляцию, обнаружены вдоль латеральной супрасильвиевой борозды, в височной, лобной и теменной коре. При их поражении нарушается пространстве
Центральный (корковый) отдел зрительного анализатора расположен в затылочной доле. В каждом участке коры по глубине сконцентрированы нейроны, которые образуют колонку, проходящую через все слои вертикально. При этом происходит функциональное объединение нейронов, выполняющих сходную функцию.
Бинокулярное зрение — восприятие окружающих предметов двумя глазами (от лат. bi — два, осulus — глаз) — обеспечивается в корковом отделе зрительного анализатора благодаря сложнейшему физиологическому механизму зрения — фузии, т. е. слиянию зрительных образов,возникающих отдельно в каждом глазу (монокулярное изображение), в единое сочетанное зрительное восприятие. Это очень тонкая функция обеспечивается двумя механизмами: согласованными движениями обоих глаз, поддерживающих постоянное направление зрительных линий на точку бификсации и слиянием изображений двух глаз в единый образ. Это позволяет обеспечить зрительной системе более высокую оценку видимых объектов (стереоскопическое зрение).
Способность зрительного анализатора определять третье измерение, телесность, стереоскопичность предметов окружающего мира, определять расстояние между предметами обусловлено одновременным зрением двумя глазами - б и н о к у л я р н ы м зрением. Бинокулярное зрение создает и другие значительные преимущества зрительному анализатору, расширяет поля зрения в горизонтальном направлении до 180 градусов, зрительные образы, полученные от двух глаз, ярче и четче вследствие суммации раздражений (острота зрения повышается), при помощи бинокулярного зрения человек более точно определяет расстояние (глазомер).
Бинокулярное зрение - это чрезмерно тонкий, сложный условно-рефлекторный комплекс. Он развивается, совершенствуется и изменяется в течение всей жизни. Огромное значение в развитии функции бинокулярного зрения играет индивидуальный опыт. Физиологический механизм бинокулярного зрения при рождении отсутствует. Несмотря на то, что глаза новорожденного рефлекторно обращены в сторону яркого раздражителя, движения их еще полностью разобщены. Лишь в возрасте 5 - 6 недель устанавливается первая бинокулярная кортикальная связь, появляются параллельные движения взора к 3 месяцам. Формирование бинокулярного зрения происходит в период от 2 месяцев до 6-10 лет и закрепляется до 15 лет. До 2 месяцев развиваются общие движения глазных яблок, укрепляются условно-рефлекторные связи между сетчаткой и движениями глаз. В акт аккомодации подключается конвергенция. В возрасте 4-5 месяцев отмечается продолжительная фиксация предмета со второго полугодия формируется фузия.
Развитие бинокулярного зрения следует рассматривать как постеленное формирование относительно устойчивой, но динамичной стереотипии нервных процессов. Уже в возрасте 2-4 месяцев у ребенка возникает функциональная взаимосвязь между обеими половинами зрительного анализатора, а также между оптическим и двигательным аппаратами, т. е. примитивное бинокулярное зрение. На ранних этапах онтогенеза бинокулярная зрительная система и ее основной саморегулирующийся оптомоторный механизм - бификсация еще недостаточно устойчивы и сравнительно легко трансформируются под влиянием неблагоприятных факторов внешней и внутренней среды. Это обусловливает возможность и создает потенциальные условия для возникновения содружественного косоглазия. Чем старше ребенок, тем устойчивее бинокулярная зрительная система и тем труднее вывести ее из состояния стереотипии.
Механизм нормального бинокулярного зрения можно представить в виде замкнутой системы врожденных и приобретенных, афферентных и эфферентных нервных связей между:
v оптическим рецептором каждого глаза и соответствующим зрительным центрам;
v кортикальными центрами обеих половин зрительного анализатора;
v зрительными центрами и корковыми центрами движений глаз;
v корковыми и подкорковыми центрами глазодвигательных мышц и самими мышцами.
Для формирорвания бинокулярного зрения необходимы определенные условия:
· Острота зрения на каждый глаз не ниже 0,3 – 0,4;
· Полный объем движений глазных яблок;
· Параллельное положение глазных яблок при взгляде вдаль;
· Соответствующая конвергенция при взгляде с близкого расстояния;
· Наличие изейконии;
· Способность к фузии;
· Попадание изображения на корреспондирующие точки сетчатки.
Стимулом к бинокулярной фиксации объекта служит постоянная тенденция зрительной системы к преодолению диплопии, к одиночному видению.
Это свойство глаз уже давно обратило на себя внимание исследователей и вызвало к жизни теорию корреспонденции сетчаток (Muller, 1826; Hering, 1868). Суть ее состоит в том, что одиночное восприятие наблюдаемого объекта возможно только при условии одновременного раздражения центральных ямок сетчаток или точек сетчаток, удаленных от центральных ямок на одинаковое расстояние в одном и том же направлении. Это идентичные, соответственные или корреспондирующие точки сетчаток. Если изображение объекта попадает на неидентичные, несоответственные или диспаратные точки сетчаток, то возникает двоение. Таким образом, ядром теории корреспонденции сетчаток служит положение о наличии в зрительной системе функционально спаренных ретино-кортикальных элементов, представляющих оба глаза.
Свойство корреспонденции сетчаток определяется как особенностями морфологического строения зрительного анализатора, так и различием задач, осуществляемых в едином зрительном акте центральным и нецентральным (периферическим) зрением.
Цветовое зрение. Теории цветоощущения. Виды цветовой слепоты.
Цветовое зрение. Весь видимый нами спектр электромагнитных излучений заключен между коротковолновым (длина волны от 400 нм) излучением, которое мы называем фиолетовым цветом, и длинноволновым излучением (длина волны до 700 нм), называемым красным цветом. Остальные цвета видимого спектра (синий, зеленый, желтый, оранжевый) имеют промежуточные значения длины волны. Смешение лучей всех цветов дает белый цвет. Он может быть получен и при смешении двух так называемых парных дополнительных цветов: красного и синего, желтого и синего. Если произвести смешение трех основных цветов — красного, зеленого и синего, то могут быть получены любые цвета.
Теории цветоощущения. Наибольшим признанием пользуется трехкомпонентная теория (Г. Гельмгольц), согласно которой цветовое восприятие обеспечивается тремя типами колбочек с различной цветовой чувствительностью. Одни из них чувствительны к красному цвету, другие — к зеленому, а третьи — к синему. Всякий цвет оказывает действие на все три цветоощущающих элемента, но в разной степени. Эта теория прямо подтверждена в опытах, где микроспектрофотометром измеряли поглощение излучений с разной длиной волны у одиночных колбочек сетчатки человека.
Согласно другой теории, предложенной Э. Герингом, в колбочках есть вещества, чувствительные к бело-черному, красно-зеленому и желто-синему излучениям. В опытах, где микроэлектродом отводили импульсы ганглиозных клеток сетчатки животных при освещении монохроматическим светом, обнаружили, что разряды большинства нейронов (доминаторов) возникают при действии любого цвета. В других ганглиозных клетках (модуляторах) импульсы возникают при освещении только одним цветом. Выявлено 7 типов модуляторов, оптимально реагирующих на свет с разной длиной волны (от 400 до 600 нм).
В сетчатке и зрительных центрах найдено много так называемых цветооппонентных нейронов. Действие на глаз излучений в какой-то части спектра их возбуждает, а в других частях спектра — тормозит. Считают, что такие нейроны наиболее эффективно кодируют информацию о цвете.
Последовательные цветовые образы. Если долго смотреть на окрашенный предмет, а затем перевести взор на белую бумагу, то тот же предмет виден окрашенным в дополнительный цвет. Причина этого явления в цветовой адаптации, т. е. снижении чувствительности к этому цвету. Поэтому из белого света как бы вычитается тот, который действовал на глаз до этого, и возникает ощущение дополнительного цвета.
Цветовая слепота. Частичная цветовая слепота была описана в конце XVIII в. Д. Дальтоном, который сам ею страдал (поэтому аномалию цветовосприятия назвали дальтонизмом). Дальтонизм встречается у 8 % мужчин и намного реже у женщин: возникновение его связывают с отсутствием определенных генов в половой непарной у мужчин Х-хромосоме. Для диагностики дальтонизма, важной при профессиональном отборе, используют полихроматические таблицы. Люди, страдающие этим заболеванием, не могут быть полноценными водителями транспорта, так как они не могут различать цвет огней светофоров и дорожных знаков. Существует три разновидности частичной цветовой слепоты: протанопия, дейтеранопия и тританопия. Каждая из них характеризуется отсутствием восприятия одного из трех основных цветов.
Люди, страдающие протанопией («краснослепые»), не воспринимают красного цвета, сине-голубые лучи кажутся им бесцветными. Лица, страдающие дейтеранопией («зеленослепые»), не отличают зеленые цвета от темно-красных и голубых. При тританопии — редко встречающейся аномалии цветового зрения, не воспринимаются лучи синего и фиолетового цвета.
Все перечисленные виды частичной цветовой слепоты хорошо объясняются трехкомпонентной теорией цветоощущения. Каждый вид этой слепоты — результат отсутствия одного из трех колбочковых цветовоспринимающих веществ. Встречается и полная цветовая слепота — ахромазия, при которой в результате поражения колбочкового аппарата сетчатки человек видит все предметы лишь в разных оттенках серого
Острота зрения, поле зрения. Методы определения остроты и полей зрения.
Острота зрения.
Остротой зрения называется максимальная способность глаза различать отдельные детали объектов.
Остроту зрения определяют по наименьшему расстоянию между двумя точками, которые глаз различает, т. е. видит отдельно, а не слитно. Нормальный глаз различает две точки, видимые под углом в 1'. Максимальную остроту зрения имеет желтое пятно. К периферии от него острота зрения намного ниже (рис. 14.11). Острота зрения измеряется при помощи специальных таблиц, которые состоят из нескольких рядов букв или незамкнутых окружностей различной величины. Острота зрения, определенная по таблице, выражается обычно в относительных величинах, причем нормальная острота принимается за единицу. Встречаются люди, обладающие сверхостротой зрения (visus более 2).
Методика исследования остроты зрения следующая. Пациент сидит лицом к таблице (экрану) на расстоянии 5 метров от него. Один глаз прикрыт непрозрачным щитком или ладонью. Пациенту показывают и просят назвать знаки, соответствующие остроте зрения 1,0. Если он все их называет верно, то показывают более мелкие знаки. Так продолжают до тех пор, пока обследуемый не начнет ошибаться. Если пациент ошибался уже в знаках соответствующих остроте зрения 1,0, то показывают более крупные знаки, следующие за ними.
Остроту зрения оценивают по ряду, в котором пациент, правильно называет все знаки. При исследовании по таблицам допускается 1 ошибка в VII-X строчках. При использовании проекторов знаков расстояние до экрана можно варьировать от 3 до 6 метров, а пациент находится рядом с прибором, и ошибок в ответах не допускается.
Таким образом, можно проверить остроту зрения от 0,1 до 1,0-2,0.
Например: Vis OD = 0,7
Vis OS = 1,0.
Исследование остроты зрения ниже 0,1 проводится разными способами в зависимости от снижения зрительных функций:
1. Определение расстояния, с которого пациент читает оптотипы I ряда. Попросить пациента подойти к таблице и определить расстояние, с которого он видит первую строчку. Рассчитать Vis по формуле Снеллена-Дондерса
![]() ,
,
где ![]() -
расстояние, с которого пациент видит
первую строчку,
-
расстояние, с которого пациент видит
первую строчку,
![]() -
50 м, т.е. расстояние, с которого оптотипы
первой строчки видны под углом зрения
в 1 минуту.
-
50 м, т.е. расстояние, с которого оптотипы
первой строчки видны под углом зрения
в 1 минуту.
2. Проверка способности считать пальцы. Пациенту демонстрируют пальцы на темном фоне (толщина пальцев приравнивается к толщине оптотипов первого ряда) и определяют расстояние, с которого он правильно их считает. Vis рассчитывают по формуле Снеллена-Дондерса.
3. Определение остроты зрения, используя оптотипы Поляка (при Vis=0,09–0,01). Они представляют собой набор кольцевых и линейных оптотипов (рис.25). Их показывают пациенту с разных расстояний, обозначенных под каждым тестом, и просят назвать направление линий или разрыва кольца. Каждому расстоянию соответствует определенная острота зрения.

Рис. 25. Оптотипы Б.Л.Поляка для измерения остроты зрения ниже 0,1
4. При Vis<0,005 (т.е. счет пальцев с расстояния ближе 25 см) указывают расстояние, с которого пациент может правильно сосчитать пальцы.
Например: Vis = счет пальцев с 5 см.
5. Исследование способности различать направленное движение ладони у самого лица (в вертикальном и горизонтальном направлении). В случае, если направление движения определено верно: Vis=движению руки у лица.
6. При отсутствии предметного зрения, проверяют светопроек-цию при помощи направления пучка света от офтальмоскопа в зрачок пациента с четырех сторон.
При правильном определении пациентом всех четырех направлений:
![]()
![]()
![]() ,
т.е. острота зрения равна светоощущению
с правильной проекцией света.
,
т.е. острота зрения равна светоощущению
с правильной проекцией света.
При неправильном определении пациентом хотя бы одного направления:
![]()
![]() ,
т.е. острота зрения равна светоощущению
с неправильной проекцией света.
,
т.е. острота зрения равна светоощущению
с неправильной проекцией света.
Если
пациент света не видит: ![]() .
.
К объективным методам исследования остроты зрения относятся нистагмовизометрия, электро-энцефалографический и условно-рефлекторный методы. Их обычно применяют в экспертных случаях при нежелании пациента правильно оценивать свои ощущения (симуляция), в раннем детском возрасте, при психических заболеваниях.
Нистагмовизометрия – метод, основанный на регистрации непроизвольных ритмических движений глазного яблока, т.е. нистагма. Оптокинетический нистагм (ОКН) возникает при наблюдении ряда однородных объектов, быстро перемещающихся в поле зрения. Он может быть вызван у всех людей за исключением тех случаев, когда острота зрения резко снижена, отмечается сужение поля зрения, а также при заболеваниях центральной нервной системы. Таким образом, ОКН возникает в тех случаях, когда глаз различает отдельно движущиеся предметы. Это и было положено в основу метода нистагмовизометрии.
В настоящее время наиболее широкое применение получил прибор «Малыш».
Он представляет собой прямоугольный корпус с экраном 10х10 см. Тест-объекты – движущиеся в горизонтальном или вертикальном направлении чередующиеся светлые и темные полосы разной ширины (от 2 до 50 мм). Пациент может находиться от экрана на расстоянии от 0,5 до 3,3 метра.
Исследование начинают с минимальной ширины полосы (2 мм) и максимального расстояния (3,3 м). Если у пациента ОКН отсутствует, увеличивают ширину полосы и, в случае необходимости, уменьшают расстояние. При появлении ОКН снимают на цифровом табло показания остроты зрения, которые рассчитаны на основании данных ширины полосы и расстояния до экрана. Этот метод можно применять при остроте зрения 0,01-1,0.
Электро-энцефалографический метод основан на изменении биопотенциалов (a-ритма) в затылочных долях человеческого мозга. В ситуациях, когда человек не фиксирует взгляд, отмечаются высокие пики a-ритма, при появлении фиксации они снижаются.
Перед началом исследования пациента просят смотреть на световой фон без знаков и начинают запись фоновой электро-энцефалограммы (ЭЭГ) от затылочных долей головного мозга. Затем на этом фоне показывают тесты определенной величины. Если пациент видит знаки, то биопотенциалы меняют свой характер, что отражается на ЭЭГ. На основании этих данных определяют остроту зрения.
Поле зрения.
Если фиксировать взглядом небольшой предмет, то его изображение проецируется на желтое пятно сетчатки. В этом случае мы видим предмет центральным зрением. Его угловой размер у человека 1,5—2°. Предметы, изображения которых падают на остальные места сетчатки, воспринимаются периферическим зрением. Пространство, видимое глазом при фиксации взгляда в одной точке, называется полем зрения. Измерение границы поля зрения производят периметром. Границы поля зрения для бесцветных предметов составляют книзу 70°, кверху — 60°, внутрь — 60° и кнаружи — 90°. Поля зрения обоих глаз у человека частично совпадают, что имеет большое значение для восприятия глубины пространства. Поля зрения для различных цветов неодинаковы и меньше, чем для черно-белых объектов.
Поле зрения измеряется в градусах, в норме его границы имеют следующие значения:
кверху – 55 градусов;
кнаружи – 90 градусов;
книзу – 65 градусов;
кнутри – 60 градусов.
Стоит отметить, что отличие на 5-10 градусов от данных значений принято считать нормой.
Два метода оценки поля зрения:
периметрия;
кампиметрия.
Контрольный способ
Это наиболее простой и достаточно неточный метод, с помощью которого выявляются сильные нарушения. Доктор (с заранее известным полем зрения, которое должно иметь нормальные границы) и пациент усаживаются друг напротив друга на расстоянии около полуметра. Врач закрывает правый глаз, исследуемый – левый (для исследования правого глаза пациента).
Затем доктор посередине между ним и пациентом проводит пальцем или белым предметом (например, карандашом) от периферии к центру. Когда больной начинает видеть объект, он говорит «да», когда предмет исчезает из поля видения, он говорит «нет».
Исследование можно проводить в четырех направлениях (медиальном, латеральном, верхнем и нижнем), но чаще проводят в восьми (четыре основные и 4 косые направления).
Если границы периферического зрения совпадают с таковыми врача, то они имеют нормальные значения. Этот метод называется контрольным, т.к. зрение врача является контролем зрения пациента.
Основным недостатком данного способа оценки периферического зрения является его неточность. Границы поля зрения приблизительны и не выражаются числовыми значениями. Такой метод наиболее часто применяется у лежачих пациентов, когда затруднительно использование различных приборов.
Инструментальный способ
Для более точного определения границ периферического зрения можно пользоваться специальными приборами – периметрами (для определения границ поля зрения на шарообразной поверхности) и кампиметрами (для исследования центрального участка поля зрения – проводится на плоскостной поверхности).
Наиболее популярны периметры Ферстера, сферические и проекционные периметры. В настоящее время большая роль принадлежит компьютерным периметрам.
Морфофункциональная организация спинного мозга. Нейронная организация сегментов спинного мозга. Функции задних и передних корешков сегментов спинного мозга. Закон Белла-Мажанди.
Морфофункциональная организация спинного мозга
Спинной мозг – наиболее древний отдел ЦНС позвоночных. Он уже есть у ланцетника, наиболее примитивного представителя хордовых.
Спинной мозг – каудальный отдел ЦНС. Он помещается в позвоночном канале и имеет неодинаковую длину у разных представителей позвоночных.
У человека в каудальном отделе позвоночного канала собираются корешки каудальных отделов спинного мозга, образуя так называемый конский хвост.
Спинной мозг характеризуется сегментарным строением. В спинном мозге выделяют шейный, грудной, поясничный, крестцовый и копчиковый отделы. Каждый отдел состоит из нескольких сегментов. Шейный отдел включает 8 сегментов (С1 – С8), грудной – 12 (Th1 – Th12), поясничный – 5 (L1 – L5), крестцовый – 5 (S1 – S5) и копчиковый – 1-3 (Со1 – Co3). От каждого сегмента отходят две пары корешков, которые соответствуют одному из позвонков и покидают позвоночный канал через отверстие между ними.
Различают дорсальные (задние) и вентральные (передние) корешки. Дорсальные корешки образованы центральными аксонами первичных афферентных нейронов, тела которых лежат в спинномозговых ганглиях. Вентральные корешки сформированы аксонами α- и γ-мотонейронов и немиелинизированными волокнами нейронов вегетативной нервной системы.
Нейронная организация сегментов спинного мозга
Нейроны спинного мозга образуют его серое вещество в виде симметрично расположенных двух передних и двух задних рогов в шейном, поясничном и крестцовом отделах. Серое вещество распределено на ядра, вытянутые по длине спинного мозга. В грудном отделе спинной мозг имеет, помимо названных, еще и боковые рога
Спинной мозг человека содержит около 13 млн. нейронов, из них 3% — мотонейроны, а 97% — вставочные. Функционально нейроны спинного мозга можно разделить на 4 основные группы:
1) мотонейроны, или двигательные, — клетки передних рогов, аксоны которых образуют передние корешки;
2) интернейроны — нейроны, получающие информацию от спинальных ганглиев и располагающиеся в задних рогах. Эти нейроны реагируют на болевые, температурные, тактильные, вибрационные, проприоцептивные раздражения;
3) симпатические, парасимпатические нейроны расположены преимущественно в боковых рогах. Аксоны этих нейронов выходят из спинного мозга в составе передних корешков;
4) ассоциативные клетки — нейроны собственного аппарата спинного мозга, устанавливающие связи внутри и между сегментами.
Мотонейроны. Аксон мотонейрона своими терминалами иннервирует сотни мышечных волокон, образуя мотонейронную единицу. Чем меньше мышечных волокон иннервирует один аксон (т. е. чем меньше количественно мотонейронная единица), тем более дифференцированные, точные движения выполняет мышца.
Несколько мотонейронов могут иннервировать одну мышцу, в этом случае они образуют так называемый мотонейронный пул. Возбудимость мотонейронов одного пула различна, поэтому при разной интенсивности раздражения в сокращение вовлекается разное количество волокон одной мышцы. При оптимальной силе раздражения сокращаются все волокна данной мышцы; в этом случае развивается максимальное сокращение мышцы.
Интернейроны. Эти промежуточные нейроны, генерирующие импульсы с частотй до 1000 в секунду, являются фоновоактивными и имеют на своих дендритах до 500 синапсов. Функция интернейронов заключается в организации связей между структурами спинного мозга и обеспечении влияния восходящих и нисходящих путей на клетки отдельных сегментов спинного мозга. Очень важной функцией интернейронов является торможение активности нейронов, что обеспечивает сохранение направленности пути возбуждения. Возбуждение интернейронов, связанных с моторными клетками, оказывает тормозящее влияние на мышцы-антагонисты.
Нейроны симпатического отдела автономной системы. Расположены в боковых рогах сегментов грудного отдела спинного мозга. Эти нейроны являются фоновоактивными, но имеют редкую частоту импульсации (3—5 в секунду).
Нейроны парасимпатического отдела автономной системы. Локализуются в сакральном отделе спинного мозга и являются фоновоактивными.
Задние корешки образуют чувствительные афферентные пути спинного мозга, а передние — двигательные эфферентные пути (рис. 1А). Такое расположение афферентных и эфферентных волокон было установлено еще в начале XX в. и получило название закона Белла-Мажанди, причем количество афферентных волокон больше количества двигательных волокон.
Задние рога выполняют главным образом сенсорные функции и содержат нейроны, передающие сигналы в вышележащие центры, в симметричные структуры противоположной стороны либо к передним рогам спинного мозга.
В передних рогах находятся нейроны, дающие свои аксоны к мышцам. Все нисходящие пути центральной нервной системы, вызывающие двигательные реакции, заканчиваются на нейронах передних рогов. В связи с этим Шеррингтон назвал их «общим конечным путем».
Альфа- и гамма-мотонейроны спинного мозга, их функции. Нейроны боковых рогов сегментов спинного мозга, их функции.
Альфа- и гамма-мотонейроны спинного мозга, их функции
Мотонейроны спинного мозга функционально делят на α- и γ-нейроны.
α-Мотонейроны образуют прямые связи с чувствительными путями, идущими от экстрафузальных волокон мышечного веретена, имеют до 20 000 синапсов на своих дендритах и характеризуются низкой частотой импульсации (10—20 в секунду).
γ-Мотонейроны, иннервирующие интрафузальные мышечные волокна мышечного веретена, получают информацию о его состоянии через промежуточные нейроны. Сокращение интрафузального мышечного волокна не приводит к сокращению мышцы, но повышает частоту разрядов импульсов, идущих от рецепторов волокна в спинной мозг. Эти нейроны обладают высокой частотой импульсации (до 200 в секунду).
Начиная с I грудного сегмента спинного мозга и до первых поясничных сегментов, в боковых рогах серого вещества располагаются нейроны симпатического, а в крестцовых — парасимпатического отдела автономной (вегетативной) нервной системы.
Симпатические, парасимпатические нейроны расположены преимущественно в боковых рогах. Аксоны этих нейронов выходят из спинного мозга в составе передних корешков;
Нейроны симпатического отдела автономной системы. Расположены в боковых рогах сегментов грудного отдела спинного мозга. Эти нейроны являются фоновоактивными, но имеют редкую частоту импульсации (3—5 в секунду).
Нейроны парасимпатического отдела автономной системы. Локализуются в сакральном отделе спинного мозга и являются фоновоактивными.
В случае поражения боковых рогов спинного мозга исчезают кожные сосудистые рефлексы, нарушается потоотделение, наблюдаются трофические изменения кожи, ногтей. При одностороннем поражении парасимпатического отдела автономной нервной системы на уровне крестцовых отделов спинного мозга нарушений дефекации и мочеиспускания не наблюдается, так как корковая иннервация этих центров является двусторонней.