

Мякушко С.А. Порівняльна анатомія. 2019
.pdfУ міру наближення очного міхура до поверхні, ектодерма, що лежить навпроти нього, потовщується та занурюється назустріч бокалу. Це потовщення відокремлює від себе в порожнину бокала структуру, з якої пізніше буде сформований кришталик. Мезодерма, що оточує закладку ока, дає початок зовнішнім оболонкам ока: судинній, яка виконує трофічну функцію, та склері, що забезпечує опорну і захисну
функції. Судинна оболонка формує війкове тіло і райдужку. Структурні елементи ока. Шар щільної сполучної тканини або
хряща утворює зовнішню оболонку ока — склеру. На передній поверхні ока склера переходить в тонку прозору рогівку, вкриту прозорим
епітелієм (рис. 10.4). Форма рогівки є мінливою: у риб вона сплощена, у наземних і вторинноводних є опуклою. Разом з кришталиком рогівка бере участь в заломлюванні потоку світла, яке надходить в око. Зсередини до склери прилягає судинна оболонка, щільно забезпечена кровоносними судинами. Нижче розташована темна пігментна оболонка.
8 |
|
9 |
10
7
11
6
5
12
4
3
2 |
14 |
13
1
Рис. 10.4. Будова ока:
1 — війкове (циліарне) тіло, 2 — задня камера, 3 — передня камера, 4 — райдужка, 5 — рогівка, 6 — кришталик, 7 — м’яз, 8 — повіка, 9 — склера, 10 — судинна оболонка, 11 — сітківка, 12 — центральна ямка жовтої плями, 13 — зоровий нерв, 14 — склоподібне тіло
(за Linzey, 2012)
310
На межі між склерою і рогівкою краї обох оболонок утворюють кільцеву складку — райдужку, яка обмежує отвір — зіницю. Остан-
ній може розширюватися або звужуватися, завдяки наявності в райдужці радіальних і кільцевих м’язових волокон. Насичення райдужки
пігментами забезпечує різноманітне забарвлення ока.
Позаду райдужки утворюється кільцевої валик — війкове тіло, яке являє собою скупчення м’язових волокон, прикріплених до оболонки кришталика. Війкове тіло бере участь в акомодації ока, змінюючи форму кришталика і, у деяких наземних хребетних, переміщаючи його відносно сітківки. За зіницею розташоване кулеподібне (у риб)
або сплюснуте в двоопуклою лінзу прозоре тіло — кришталик. Внутрішній шар стінки ока — сітківка — щільно прилягає до
пігментної оболонки. Остання складається із світлочутливих, нервових та опорних клітин. До пігментного шару прилягає шар фоторецепторів, зовнішні членики яких мають вигляд паличок або колбочок
імістять світлочутливий пігмент. Палички і колбочки мають різні будову та функції. Палички — рецептори, які реагують на слабке освітлення, вони передають зображення в чорно-білих тонах. Палички переважають у сітківці нічних наземних тварин, в акул і глибоководних риб. Колбочки чутливі до інтенсивного освітлення, вони краще передають дрібні деталі зображення і забезпечують кольоровий зір.
Умісці виходу зорового нерва світлочутливий шар сітківки редукується, цю ділянку називають сліпою плямою. У центрі сітківки зазвичай розташовується область підвищеної оптичної чутливості — жовта пляма; в її центрі часто є ямка, де щільність фоторецепторів
інервових клітин збільшена. Внутрішня порожнина очного яблука заповнена склоподібним тілом. Усередині цього тіла у рептилій і птахів наявна структура, щільно пронизана кровоносними судинами — сосковий конус, або гребінець. Він покращує забезпечення очей поживними речовинами та киснем (також, можливо, сприяє гостроті зору). Рух очей в орбіті обумовлений скороченням чотирьох прямих
ідвох косих м’язів, які прикріплюються до стінок орбіти та склери.
Для отримання чіткого зображення існує механізм «наведення» очей на різкість — акомодація.
Акомодація у хребетних може здійснюватися по-різному (рис. 10.5). У цьому процесі беруть участь мезодермальні м’язи
311

|
|
4 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
3 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
2 |
|
|
|
|
|
|
|
|
|
|
|
|
|
1 |
|
2 |
|
|
|
|
|
|
1 |
|
|||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
6 |
|
|
|
|
|
|
|
|
|
|
|
|
|
||||
|
|
5 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
6 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||
|
|
|
|
|
|
|
|
7 |
|
|
|
|
|
|
|
|
|
|
|
||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
7 |
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
8 |
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
8 |
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
4 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
6
5
Рис. 10.5. Механізми акомодації у хребетних:
А— круглороті, Б — костисті риби, В — черепахи, Г — птахи,
Д— ссавці, 1 — стан спокою, 2 — стан акомодації, 3 — кришталик,
4 — рогівка, 5 — війковий м’яз, 6 — сфінктер райдужки, 7 — склеральні скостеніння, 8 — циліарний відросток (за Kardong, 2012)
війкового тіла та ектодермальні м’язи райдужної оболонки. У круглоротих, хрящових і кісткових риб, у личинок амфібій акомодація досягається переміщенням кришталика в середині очного яблука, у рептилій, птахів і ссавців пристосування до чіткого бачення відбувається за рахунок переміщення і зміни форми кришталика. У разі розглядання віддалених предметів кришталик сплощується, при ближньому зорі він стає округлим. Ці зміни обумовлені роботою м’яза війкового тіла. Помічено, що спокійний стан кришталика у різних хребетних може забезпечувати досягнення різних завдань: в одних
312
випадках проблему близького зору, в інших — далекого. Загалом виділяють до семи різних і незалежно сформованих варіантів акомодації в окремих групах тварин.
З органами зору хребетних пов’язані різноманітні допоміжні структури: повіки, слізні залози та ін. Повіки практично відсутні у риб, в окремих випадках біля очей дещо розростаються складки
шкіри, у деяких акул можуть бути верхня і нижня повіки.
У наземних хребетних очі завжди прикриті повіками. Їх функція полягає в захисті рогівки від висихання, а також від попадання чужорідних часточок і пилу. В амфібій, рептилій і птахів сильніше розвинена нижня повіка, у ссавців і крокодилів — верхня. Під рухливими повіками амфібій, рептилій, птахів є третя повіка — мигальна перетинка, яка прикриває око спереду назад. У змій повіки зростаються, вони дуже тонкі, тому рогівка виявляється прикритою прозорою шкірою. Слізні залози наземних хребетних розташовані в передньому
куті ока. Їх рідкий секрет змочує і промиває рогівку. У передньому куті ока знаходиться гардерова залоза, яка секретує маслянисту рі-
дину для змочування передньої поверхні очного яблука. Гардерові залози добре розвинені у земноводних, плазунів і птахів. Ссавцям вони, як правило, не властиві, за виключенням китоподібних і ластоногих, у яких секрет цих залоз захищає очі у воді, а над водою збільшує далекозорість за принципом контактної лінзи. У наземних хребетних в передньому куті ока є носослізна протока. Вона виводить надлишок слізної рідини в носову порожнину, зволожуючи слизову носа. У морських черепах і деяких морських птахів слізні залози виводять гіпертонічний розчин солі, що дає можливість їм використовувати для пиття морську воду і звільнятися від надлишку солей.
В окремих випадках спрощення або редукція очей можуть бути обумовлені відсутністю освітлення. Це властиво глибоководним рибам і деяким тваринам, які ведуть підземний спосіб життя — кротам, сліпакам, гоферам та ін. Більшість тварин мають монокулярний бічний зір, але у деяких (птахи, примати) виникає бінокулярний зір, при цьому очі зближуються на лицьовому диску.
Непарні очі хребетних розвиваються як порожнисті вирости
епіталамуса — покрівлі проміжного мозку. Вони представлені пінеальним (corpus pineale) та розташованим перед ним парієтальним
313
(corpus parapineale) органами (перший відомий також як епіфіз, другий — тім’яний орган).
Обидва органи можуть мати фоторецепторні клітини, проте в еволюційному ряду хребетних є дуже мінливими. Світлочутлива функція властива тім’яному органу в деяких риб, хоча у круглоротих і безхвостих амфібій ця функція належить епіфізу. Будова тім’яного органа значно варіює у рептилій. Наприклад, тім’яний орган гатерії має вигляд пухирчастого ока. Його передня стінка потовщена та утворює кришталик, а задня стінка сильно пігментована. У деяких безхвостих амфібій та ящірок на рівні проміжного мозку, на шкірі або під нею є світлочутлива пляма. В усіх випадках непарне око сприймає рівень освітленості, а не образ предмета. Епіфіз, втративши зорову функцію, перетворюється на залозисте утворення — шишкоподібну залозу. У ссавців вона бере участь в регуляції роботи статевих залоз і формуванні вторинних статевих ознак.
Бічна лінія. Еволюційне значення системи органів бічної лінії полягає у тому, що вона дала початок іншим сенсорним органам, зокрема органам слуху і рівноваги. Ця сейсмосенсорна система притаманна первинноводним хребетним, личинкам земноводних, амфібіям, які постійно живуть у воді; була вона і у вимерлих форм первинноводних.
В онтогенезі хребетних бічна лінія закладається в ділянці ектодерми зародка, розташованій поблизу нервової пластинки. Тут утворюються потовщені ділянки, так звані чутливі горбки. Кожна така ділянка несе клітини двох типів: опорні — довгі циліндричні, що йдуть від поверхні до основи, і сенсорні — клітини грушоподібної форми. Вони займають центральне положення в ділянці, але не доходять до її основи. Сенсорні клітини на вільному кінці мають щіточку волосків,
а біля основи обплетені нервами. Скупчення чутливих клітин мають назву невромастів. Невромасти за будовою нагадують смакові цибу-
лини (рис. 10.6). Їх волоски зверху вкриті масою драглистої субстанції, яку секретують клітини невромастів; утворюється так звана купула. Купули вільних невромастів (деякі амфібії) мають видовжену циліндричну форму і виступають назовні у воду; купули канальних невромастів (риби) більш короткі, сплощеної форми. Канали заповнені рідиною з підвищеним вмістом калію і контактують із зовнішнім
314
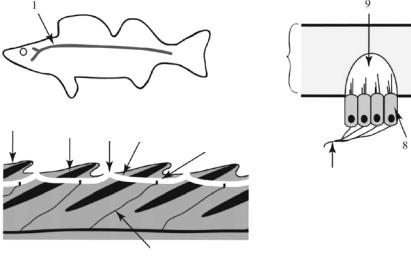
середовищем через пори. У деяких видів риб пори закриті мембраною, коливання води передаються до сенсорних клітин через ці мембрани, що нагадує передачу коливань через барабанну перетинку.
1 |
|
9 |
|
|
|
1
2
3 4 5
6 |
8 |
7
7
Рис. 10.6. Бічна лінія риб:
1 — латеральний канал бічної лінії, 2 — епідерміс, 3 — луска, 4 — пора, 5 — канал, 6 — невромаст, 7 — чутливий нерв,
8 — чутлива волоскова клітина, 9 — купула (за Hickman et al., 2012)
Органи бічної лінії являють собою шкірні канали, розташовані з боків тіла, а також на голові тварини. У круглоротих, нижчих примітивних риб і химер, у земноводних вони лежать на поверхні шкіри або занурюються в неглибокі ямки. У більшості риб канали занурені глибоко в шкіру, прикриті лускою і вздовж усього каналу мають численні виходи крізь отвори в лусочках. Головні канали бічної лінії розташовуються з боків вздовж тіла, на голові риби бічна лінія утворює складну сітку. У деяких хрящових і кісткових риб канали являють собою відкриті жолобки або борозни. У сучасних земноводних невромасти розміщуються в шкірі більш-менш ізольовано, але лінійно, як у круглоротих.
Органи бічної лінії сприймають легкі рухи і коливання води високої частоти, в основному поблизу джерела коливань. Інформація, що
315
надходить від бічної лінії, допомагає тварині в орієнтації, пошуку їжу, уникненні перешкод, запобіганні хижаків та ін. Серед амфібій бічна лінія властива личинкам безхвостих і хвостатих, які мешкають у водному середовищі, а також небагатьом видам дорослих хвостатих. У більшості безхвостих після метаморфозу бічна лінія втрачається. У амніот, які вторинно повернулися у водне середовище, бічна лінія не розвивається.
Клітини бічної лінії можуть брати участь в електрорецепції (пластинозяброві, ганоїдні, сомові). Серед електрорецепторних органів розрізняють ампульовані та горбикові. Ампульовані органи зустрічаються у всіх морських і прісноводних електрочутливих риб; горбикові поширені тільки серед прісноводних слабоелектричних костистих риб (наприклад, для орієнтації в каламутній воді, де
можливості зору обмежені).
Органи рівноваги і слуху. Через первинноводність хребетних тварин, утворенню органа слуху, тобто органа сприймаючого звукові подразники, передувала стадія, коли головною функцією відповідних структур було відчуття рівноваги. У всіх хребетних ця первинна функція вуха залишається майже незмінною. Функція слуху з’явилася пізніше, з виходом хребетних на суходіл і поступово вдосконалювалася в еволюційному ряду хребетних. Комплекс структур органів рівноваги і слуху в найрозвинутішому вигляді (у ссавців) складається з трьох компонентів: внутрішнього вуха, середнього і зовнішнього.
Упервинноводних він представлений лише внутрішнім вухом.
Укруглоротих і риб орган слуху утворений капсулами внутрішнього вуха, які закладаються з боків голови зародка у вигляді парних потовщень ектодерми. Кожна з них перетворюється спочатку на ямку, а потім на пухирець, що з’єднаний з поверхнею каналом. Пізніше він відшнуровується від ектодерми. Слуховий пухирець перехватом розділяється на два відділи: з верхнього формується овальний мішечок і з’єднані з ним півколові канали — орган рівноваги (вестибуляр-
ний апарат). Нижній відділ перетворюється на круглий мішечок, від якого відходить порожнистий виступ — лагена (lagena). Остання
увищих хребетних утворює завитку — орган слуху. Епітелій, що вистилає перетинчастий лабіринт, на окремих ділянках круглого та овального мішечка, а також в півколових каналах має чутливі клі-
316
тини, забезпечені пружними волосками. Внутрішня порожнина перетинчастого лабіринту заповнена ендолімфою. У ендолімфі мішечків,
півколових каналів і лагени зважені кристалічні включення карбонату кальцію — великі (оттоліти) і дрібні (оттоконіі). Форма от-
толітів видоспецифічна і дає можливість, наприклад, у риб визначити їх систематичну приналежність. Зміна положення голови спричинює переміщення ендолімфи та отолітів, які подразнюють при цьому чутливі клітини. Перетинчастий лабіринт, таким чином, функціонує як орган рівноваги. Звукові хвилі, які обумовлюють коливання ендолімфи і включень, що знаходяться в ній, також викликають подразнення чутливих клітин. Слід зазначити, що в процесі еволюції щелепноротих від риб до вищих ссавців будова вестибулярного апарату принципово не змінюється. Можливо, це пояснюється сталістю характеристик гравітаційного поля Землі, стабільних за напрямком і величиною та незалежних від середовища мешкання тварин.
Для більшості тварин характерна наявність у вестибулярному апараті трьох півколових каналів. Вони лежать в трьох взаємно перпендикулярних площинах у відповідності з трьома просторовими вимірами. Часто обидва вертикальних канали починаються від верхньої сторони овального мішечка спільною ніжкою. Кожний із півколових каналів має на одному із своїх кінців розширення, яке називається — ампула, в кожній із ампул є чутливий гребінь. У міног наявні лише два канали і такий стан, ймовірно, є вихідним. Вони розташовані у взаємно перпендикулярних площинах і сприймають кутові прискорення, які відчуває голова, щодо всіх трьох осей. Вважають, що в кожному відносно невеликому за діаметром півколовому каналі міноги, ендолімфа не тільки знаходиться в круговому русі, але й підганяється джгутиковим апаратом. Міксинам властивий тільки один канал. Проте, скоріше за все, така ситуація є вторинною, тобто є результатом зростання двох каналів.
Спеціалізований слуховий рецептор у внутрішньому вусі риб відсутній. Риби, земноводні та плазуни сприймають звуки всією поверхнею тіла, оскільки позбавлені зовнішнього вуха. Органами, що сприймають коливання середовища, у риб і личинок земноводних є внутрішнє вухо, бічна лінія та клітини шкірного аналізатора. Слухову функцію у внутрішньому вусі виконують окремі волоскові клітини
317
круглого мішечка і лагени — рецептор тиску. Вони вловлюють звуки, віддалені на великі відстані, а бічна лінія вловлює звуки поблизу їх джерела. Інтенсивні звуки може сприймати шкірний аналізатор. Виявлено, що хрящові риби сприймають поверхнею тіла звукові коливання низьких частот.
Усприйнятті звуків кістковими рибами беруть участь система порожнин в черепі та плавальний міхур. В оселедцеподібних звукові коливання з поверхні тіла передає трубчастий відросток плавального міхура, який контактує з перетинчастим лабіринтом. У коропоподіб-
них механізм інший: плавальний міхур пов’язаний з перетинчастим лабіринтом системою кісточок — похідних відростків хребців (веберів апарат). Сполучення плавального міхура з внутрішнім вухом здійснюється за допомогою чотирьох пар веберових кісточок. Рухомо зчленовані веберові кісточки функціонально являють собою аналог системи кісточок середнього вуха ссавців, а плавальний міхур — функціональний аналог барабанної перетинки.
В умовах наземного середовища характеристики звукопередачі інші. Швидкість поширення звукової хвилі в повітрі менше, ніж
уводі, звуки швидко розсіюються і загасають. У повітряному середовищі механічні звуки, які поширені серед риб, стають малоефективними. Тому системи сприйняття і створення звуків у наземних хребетних суттєво змінюються.
Рецепторний апарат внутрішнього вуха амфібій характеризується появою так званих сосочків. Між овальним і круглим мішечками розташовується амфібійний сосочок — новоутворення земноводних, відсутнє в інших наземних хребетних. За його участю тварини сприймають звуки в досить вузькому діапазоні: 200–940 Гц. Вважають, що амфібійний сосочок також бере участь у системі водного сприйняття звуків. Інший сосочок — базилярний — розташовується в круглому мішечку поблизу лагени. Він сприймає звуки в діапазоні 3–4 Гц — 1 кГц. Передача звуків на рецепторний апарат внутрішнього вуха здійснюється в амфібій двома шляхами: водним (через лімфатичні мішки, венозну систему та ендолімфу внутрішнього вуха) і повітряним (через середнє вухо).
Урептилій амфібійний сосочок відсутній. У круглому мішечку помітний виріст, який отримав назву — кохлеарний канал (або
318
завитка). Тут сконцентровані волоскові клітини базилярного сосочка і лагени. У внутрішньому вусі птахів, як і у рептилій, рецепторним органом є базилярний сосочок. На дистальному кінці завитки розташована лагена. Волоскові клітини птахів різноманітні — відрізняються будовою, розташуванням та іннервацією. Завитка має збільшені розміри овального і круглого вікон. На базилярній мембрані у птахів, окрім сосочка, розвивається витягнутий у довжину кортієв орган, який складається із рецепторних і опорних клітин, покритих (як і сосочок плазунів) мембраною.
У внутрішньому вусі ссавців завитка спірально закручена в 1–5 оборотів і повністю ізольована від вестибулярного апарату (в однопрохідних вона має 0,5 обороту і нагадує завитку птахів). В завитці ссавців, крім однопрохідних, зникає макула лагени і весь рецептор, представлений базилярним сосочком, видозміненим у справжній кортієв орган ссавців, розміщений на базилярній мембрані (рис. 10.7). Специфічною рисою ссавців є постійна кількість рецепторних клітин і однаковий характер їх розташування.
Середнє вухо є характерним утворенням наземних хребетних. Воно забезпечене барабанною перетинкою і різною кількістю слухових кісточок. На середнє вухо перетворюються IV зяброва щілина — бризкальце і верхній елемент під’язикової дуги — підвісок (гіомандібуляре). Зовні порожнина середнього вуха обмежена тонкою мембраною — барабанною перетинкою. Остання утворена листком сполучної тканини і прикрита з поверхні шкірою (в амфібій, багатьох ящірок). Через широкий отвір або через євстахієву трубу ця
порожнина з’єднується з глоткою. Гіомандібуляре втрачає роль підвіска щелеп і перетворюється на стремінце (stapes).
Розширений у вигляді підошви кінець стремінця впирається в овальне вікно внутрішнього вуха і передає коливання барабанної перетинки на систему: перилімфатична протока — базилярна мем-
брана — лагена. У ссавців в середньому вусі формується три слухові кісточки — молоточок (malleus), коваделко (incus) і стремінце,
гомологи відповідно квадратної, зчленівної кісток і підвіска. Вважають, що система кісточок не тільки ефективніше передає і підсилює коливання, але й оберігає внутрішнє вухо від навантажень при швидких змінах інтенсивності звуків.
319