

Мякушко С.А. Порівняльна анатомія. 2019
.pdf6.Як відбувається внутрішнє запліднення, властиве деяким групам риб?
7.Яке походження і значення має клоака в деяких групах хребетних?
8.Охарактеризуйте органи, які забезпечують внутрішнє запліднення в різних рядах хребетних тварин.
9.У чому полягають зміни статевої системи у первинноводних хребетних?
10.Назвіть відмінності між яйцекладінням, яйцеживородінням
іживородінням.
11.Чим обумовлена поява вторинних статевих протоків кісткових риб?
12.Охарактеризуйте будову яєць птахів.
13.Порівняйте будову яєць рептилій і птахів.
14.Чим обумовлено функціонування у птахів одного яєчника та одного яйцепроводу?
15.Де відбувається запліднення в різних групах хребетних тварин?
16.Чим обумовлено відокремлення різних типів маток у ссавців?
17.Які функції виконує плацента ссавців?
18.Які типи плацент властиві представникам різних груп ссавців?
270
РОЗДІЛ 8 ЕНДОКРИННА СИСТЕМА
Ендокринні залози та їх гормони знаходяться у тісному зв’язку з нервовою системою, утворюючи загальний інтеграційний механізм регуляції. Регулюючий вплив ЦНС на фізіологічну активність залоз внутрішньої секреції здійснюється через гіпоталамус.
Залози внутрішньої секреції не мають спеціальних вивідних проток в органи. Секрети ендокринних залоз — гормони — надходять у кровоносне русло та з током крові розносяться по всьому тілу. Для більшості ендокринних залоз виділення гормонів є єдиною їх специфічною функцією (паращитоподібна, щитоподібні, зобна, надниркові залози, гіпофіз), для інших виділення гормонів є додатковою функцією поряд з основною (підшлункова залоза, гонади). Залози внутрішньої
секреції топографічно розташовуються в різних ділянках тіла тварини. Щитоподібна залоза (glandula thyroidea) в ембріогенезі з’яв-
ляється як серединне карманоподібне випинання вентральної стінки глотки. Вона наявна в усіх хребетних. У круглоротих тканина залози розкидана вздовж глотки, в риб і деяких рептилій є компактним органом. В акул вона розташована між гілками нижньої щелепи попереду черевної аорти; у костистих риб охоплює черевну аорту в області передніх зябрових дуг; у земноводних розміщена поблизу під’язикового апарату; в рептилій — під трахеєю; у птахів — в основі бронхів; у ссавців — поблизу гортанних хрящів (рис. 8.1).
Найчастіше є непарним органом ендокринної системи, проте складається з двох часток, перешийка та рудиментарної пірамідної частки. Щитоподібна залоза має фіброзну оболонку, яка формує внутрішній та зовнішній листки, між ними знаходиться жирова клітковина, де проходять судини та гілки нервів. У ссавців спереду залозу оточують м’язи, а задня частина примикає до перснеподібного хряща та кілець трахеї. Завдяки зв’язку з гортанню щитоподібна залоза піднімається та опускається у разі ковтання, зміщується в боки під час поворотів голови. Іннервується симпатичними, парасимпатичними та соматичними нервовими гілками.
Вважають, що щитоподібна залоза хребетних є гомологом ендостиля ланцетника та покривників. Ендостиль являє собою борозну,
271

6
5
4
3
2 |
7 |
|
1
8
9
10
12
11
Рис. 8.1. Щитоподібна залоза ссавців:
1 — залоза, 2 — паращитоподібна залоза, 3 — перснеподібний хрящ, 4 — перснечерпакуватий м’яз, 5 — щитоподібний хрящ, 6 — надгортанник, 7 — рідина у порожнині, 8 — головні клітини, 9 — капіляр,
10 — парафолікулярна клітина, 11 — нерв, 12 — судина (за Kardong, 2012)
яка проходить на вентральній стінці глотки та вислана війчастим і залозистим епітелієм. В цій борозні або жолобі поряд із слизом утворюються йодисті сполуки, які вносяться у травний тракт разом з їжею. Останнє відрізняє ендостиль від щитоподібної залози, гормони якої надходять в кров’яне русло. Перехідна структура між ендостилем
272
і щитоподібною залозою хребетних має місце в круглоротих. У личинки міноги залоза закладається на вентральному боці глотки у вигляді поздовжнього жолоба, стінка якого вистелена миготливим епітелієм і містить декілька рядів залозистих клітин. Перед метаморфозом цей жолобок відшнуровується від глотки та перетворюється на пухирець, епітелій якого утворює групу окремих фолікулів, що характерні для щитоподібної залози.
Для мікроструктури залози є характерними велика кількість таких округлих фолікулів, а також розвинена сітка кровоносних судин. Порожнини фолікулів містять у колоїдному стані білок тиреоглобулін. Його молекула включає йод, тому ця речовина має назву йодтиреоглобулін. В результаті гідролізу цього білка в кров виділяються гормони тироксин і трийодтиронін. Останні беруть участь у регуляції загального тканинного та енергетичного обмінів. У птахів і ссавців ці гормони контролюють підтримання високої температури тіла, в амфібій — метаморфоз. Існують дані, що під час зимової сплячки активність залози у ссавців знижується, а в амфібій, навпаки, підвищується. Гіпофункція щитоподібної залози сповільнює обмін речовин, затримує ріст, порушує терморегуляцію. Ендокринна активність
щитовидної залози контролюється тиреотропним гормоном гіпофіза.
Паращитоподібні залози (glandula parathyroidea) в ембріогенезі закладається з черевного боку глотки одночасно зі щитоподібною у вигляді кількох часточок. Структурно вони відрізняються від щитоподібної залози характером розташування клітин. Їх паренхіма представлена різними за формою епітеліальними тяжами (трабекулами) та скупченнями клітин — паратироцитів, між якими розташовані капіляри. Залози мають власні волокнисті капсули, іннервуються від шийних гангліїв (симпатична система) і блукаючого нерва (парасимпатична). Паращитоподібні залози тварин вивчені доволі слабо. Відомо, що вони не оформлені в риб, але добре розвинені в змій і ссавців. У останніх залози представлені зазвичай двома парами, які утворюються із вентральних стінок зябрових карманів і розташовані з боків щитоподібної залози.
Гормон паращитоподібних залоз — білок паратгормон — регулює концентрацію кальцію та фосфору в тканинах і рідинах організму. Певна концентрація кальцію особливо важлива для нервово-м’язових
273

|
|
|
|
|
|
|
взаємодій. Гормон також є важли- |
|
|
|
|
|
|
|
|
вим для підтримання хімічного |
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
складу кісткової тканини: якщо не |
|
|
|
|
|
|
||||
|
|
|
|
3 |
|
|||
2 |
||||||||
|
|
|
|
|
надходить достатньої кількості фос- |
|||
|
|
|
|
|
||||
|
|
|
|
|
|
|
фору і кальцію (для утворення фос- |
|
|
|
|
|
фату кальцію) оновлення кісткової |
||||
|
|
|
4 |
|
||||
1 |
||||||||
|
|
|
|
|
|
|
тканини гальмується, а скелет де- |
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
генерує. Паращитоподібні залози |
|
5впливають також на нервову подраз-
ливість, проникність клітинних
мембран, здатність крові зсідатися.
Тимус, або загруднинна (зобна, вилочкова) залоза (thymus) має
часточкову будову та розміщується в області трахеї у передній частині грудної порожнини (рис. 8. 2). В онтогенезі хребетних розміри та значення цієї залози змінюються: вона збільшується до настання статевої зрілості, з віком зазнає інволюції
та деградує, а іноді й повністю зникає (наприклад, у ссавців). Тимус виконує імунорегуляторну, ендокринну та лімфопоетичну (виробляє лімфоцити) функції. Саме він формує Т-клітини, що є центральними регуляторами відповіді імунної системи.
З’являється у вигляді випинань зябрових кишень спинного боку глотки. У риб зачатки цієї залози закладаються поблизу зябрових мішків, у наземних хребетних тимус розвивається з меншої кількості зачатків і розташовується з боків шиї.
Гіпофіз (hypophysis) в ембріогенезі формується з кількох джерел: із проміжного мозку у вигляді вентрального виросту порожнистої пальцеподібної воронки, а також з ектодермального епітелію
первинної ротової порожнини. Цей епітелій випинається в напрямку до проміжного мозку та формує так званий гіпофізарний карман
(карман Ратке). Останній потім відшнуровується від епітелію ротової порожнини. Зв’язок з епітелієм у дорослих особин зберігається тільки в деяких риб і круглоротих.
274
Відповідно до ембріонального походження у гіпофізі виділяють дві основні частини: залозистий гіпофіз, або аденогіпофіз, який складає більшу частину залози і розвивається із кармана Ратке та
нейрогіпофіз, який розвивається із проміжного мозку.
Доведено, що гормони, які накопичуються в нейрогіпофізі, виробляються ядрами гіпоталамуса. Тіла клітин гіпоталамуса характеризуються секреторною активністю, а їх аксони передають гормони в нейрогіпофіз. Гормони нейрогіпофіза — вазопресин і окситоцин. Перший стимулює реабсорбцію води в нирках і регулює водний баланс, другий впливає на процеси, пов’язані з розмноженням: він
стимулює скорочення мускулатури матки та процес лактації. Аденогіпофіз представлений скупченнями та тяжами секретор-
них епітеліальних клітин, поділених каркасом зі сполучної тканини. В ньому виділяють дистальну, горбикову та проміжну частки; остання має тенденцію зливатися з нервовою часткою гіпофіза. Залозисті клітини розміщені в дистальній та проміжній частинах аденогіпофіза, горбикова частка позбавлена секреторних клітин. Аденогіпофіз виробляє більшість гормонів гіпофіза: гормон росту стимулює ріст скелета та мускулатури, регулює обмін жирів і вуглеводів; пролактин стимулює у ссавців секрецію молока, у деяких птахів — «пташиного молочка»; гонадотропні гормони регулюють функції яєчників і сім’яників; тиреотропний і аденокортикотропний гормони стимулюють роботу щитоподібної та надниркових залоз. Діяльність аденогіпофіза регулюється мозком через ворітну систему, розташовану на його межі з нейрогіпофізом.
Нейрогіпофіз побудований інакше, тут добре розвинена сітка кровоносних судин і знаходяться «запаси» гормонів. Раніше вважали, що гормони виробляються спеціальними клітинами з розгалуженнями, які тут розташовані. Зараз вчені дотримуються думки, що ці розгалужені клітини являють собою опорні елементи типу нейроглії в структурах ЦНС. Гормони ж виробляються не в залозі,
ає секретом клітин гіпоталамуса.
Укруглоротих нейрогіпофіз є недорозвиненим, він щільно по- в’язаний з потовщеним дном проміжного мозку; аденогіпофіз зберігає зв’язок з епітелієм ротової порожнини. У хрящових і променеперих риб обидві частини залози добре розвинені, наявний судинний мішок.
275
У дводишних риб цей мішок відсутній; нейрогіпофіз сплощений, потовщена лише його каудальна ділянка. Зберігається взаємне проникнення тканин нейрота аденогіпофіза, яке характерне для костистих риб.
В амфібій та рептилій має місце збільшення нейрогіпофіза, тенденція ця зберігається також у ссавців, а сама залоза набуває сферичної форми. Проміжна частка гіпофіза розвинута по-різному: вона відсутня у птахів, слабо виражена в ссавців, особливо комахо-
їдних, неповнозубих, хижих і рукокрилих.
Надниркові залози (glandula suprarenalis) виникають в ембріогенезі як похідні зачатків симпатичних вузлів вегетативної нервової системи, від чого у нижчих хребетних зберігають дифузний стан. У амніот ця тканина пізніше стає мозковою речовиною компактних тіл цих залоз. Кіркова речовина залоз закладається ембріонально в мезодермі міжниркової ділянки.
Парні надниркові залози знаходяться у рептилій, птахів, ссавців зазвичай біля переднього верхнього краю нирки. У риб та амфібій їх ділянки розкидані вздовж нирок. У круглоротих і деяких риб клітини мозкової та кіркової речовини можуть бути розкиданими між нирками, навколо них, вдовж великих кровоносних судин; у акул компоненти двох шарів відособлені між собою; в амфібій, плазунів і птахів обидва компоненти чітко не поділені; тільки в ссавців надниркові залози являють собою компактні тіла з чітким пошаровим розмежуванням.
Клітини кіркового шару секретують стероїдні гормони, які по-різ- ному впливають на функції організму. Сукупність гормонів цього шару найчастіше поділяють на дві групи — глюкокортикоїди та мінералокортикоїди. Окремі гормони кіркової речовини беруть участь у регуляції водно-сольового обміну, що вказує на можливий зв’язок між кірковою речовиною та нирками. В цілому гормони кори надниркових залоз дозволяють організму витримувати тривалі зовнішні стреси, а гормони мозкової речовини — короткочасні критичні ситуації.
Мозкова речовина надниркових залоз складається з видозмінених нервових клітин — постгангліонарних нейронів. В ембріогенезі вони утворюються із клітин, які мігрують з нервового гребеня вздовж вісцеральних нервів, тобто мають таке ж походження, як і постгангліонарні нейрони симпатичної нервової системи. Клітини мозкового шару у разі відповідної стимуляції виділяють у кров дві хімічно
276
близьких речовини: адреналін і норадреналін. Відомо, що ці речовини виділяються також постгангліонарними волокнами симпатичної нервової системи, проте клітини мозкового шару надниркових
залоз виділяють їх у значно більшій кількості.
Епіфіз, або шишкоподібне тіло (glandula pinealis), розташований у глибині головного мозку. Основними функціями є регуляція циркадних (добових) біологічних ритмів, ендокринних функцій, метаболізму, пристосування організму до змінних умов освітленості. Гормони епі-
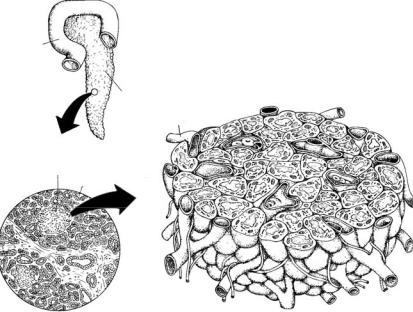
фізу також певний час гальмують розвиток і функцію статевих залоз. Підшлункова залоза (pancreas) належить до групи залоз мі-
шаної секреції (рис. 8.3). В ембріогенезі вона відгалужується від середньої частини кишечнику. Більша частина тканини цієї залози виробляє травні ферменти, що надходять протоками у травну трубку.
Реалізацію ендокринної функції забезпечують ділянки залозистої тканини — острівці Лангерганса. Описано два типи клітин острівців,
які продукують різні гормони. У круглоротих, риб і деяких ссавців вони розкидані вздовж кишки. Гормон залозистих клітин — інсулін — виділяється безпосередньо у кров. Він бере участь у регуляції вуглеводного обміну, знижуючи рівень глюкози в крові та стимулюючи синтез і накопичення вуглеводів у печінці. Разом з інсуліном залозисті клітини підшлункової залози виробляють інший гормон — глюкагон. Він сприяє розщепленню зарезервованого в печінці глікогену, що супроводжується підвищенням рівня глюкози в крові.
Гастроінтестинальні гормони утворюються неспеціалізованими клітинами. Так, у пілоричній частині шлунка виробляється гормон гастрин, який стимулює секрецію соляної кислоти. В передній частині тонкої кишки виробляється гормон, який відповідає за подачу в кишечник соку підшлункової залози. Інший кишковий гормон — панкреохолецистокінін, стимулює утворення ферментів підшлункової залози та спорожнення жовчного міхура.
Гонади також характеризуються ендокринною функцією, яка, своєю чергою, знаходиться під впливом гормонів гіпофіза. Ендокринна діяльність гонад відіграє основну роль у формуванні вторинних статевих ознак, проявах статевого диморфізму, в забарвленні шкіри тварин, характері сезонного оперення птахів і волосяного покриву ссавців, бере участь у регуляції процесів, пов’язаних з розмноженням.
277
1
2
5
3
4
Рис. 8.3. Підшлункова залоза ссавців:
1 — дванадцятипала кишка, 2 — тіло залози, 3 — острівці Лангерганса, 4 — екзокринна частина залози, 5 — капіляр (за Kardong, 2012)
Статеві гормони, які виробляються гонадами, за своєю природою є стероїдами. Стероїдні гормони гонад за хімічною структурою дуже подібні до гормонів кори надниркових залоз. Із андрогенів відомі два гормони — тестостерон і андростендіон, із естрогенів —
естрадіол і естрон.
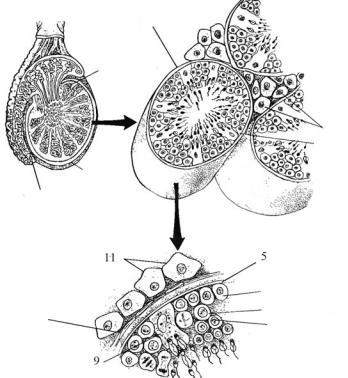
Ендокринну функцію в гонадах виконують особливі інтерстиціальні клітини, які походять із мезенхіми (рис. 8.4). Вони не вияв-
лені у нижчих хордових, проте наявні в усіх хребетних. Інтерстиціальні клітини не входять до складу сім’яних канальців чи міхурців,
а розміщені в сполучній тканині у проміжках між канальцями і пухирцями, тобто — інтерстиціально. Клітини Сертолі мають ана-
логічне походження, але розміщені, на відміну від перших, у стінках канальців разом із статевими клітинами. Вони не виробляють гормонів, проте їх секрет зв’язує андрогени, цим вони підтримують їх високу концентрацію в сім’яниках.
278
3
3
4
5
1
2
11 |
|
5 |
6
10
7
8
Рис. 8.4. Мікроструктура сім’яника ссавця:
1 — сім’яник, 2 — придаток сім’яника, 3 — сім’яний каналець, 4 — інтерстиціальні клітини, 5 — базальна пластинка, 6 — сперматогоній,
7 — первинний сперматоцит, 8 — вторинний сперматоцит, 9 — клітини Сертолі, 10 — сполучнотканинна капсула (за Kardong, 2012)
Основним джерелом естрогенів в яєчниках є фолікулярні клітини; частину естрогенів можуть виробляти також інтерстиціальні клітини, які в невеликих кількостях наявні в яєчниках. У ссавців, при виході яйцеклітини із яєчника, фолікул, що вміщав цю яйцеклітину, заповнюється гіпертрофованими фолікулярними клітинами, які утворюють жовте тіло. Останнє секретує важливий стероїдний гормон ссавців — прогестерон, який готує епітелій матки до імплантації заплідненої яйцеклітини і стимулює розвиток плаценти. Утворення жовтого тіла описано також у пластинозябрових риб.
279