

Вызванное и спонтанное высвобождение медиаторов из пресинапса
В нейронах происходит генерация одного или нескольких потенциалов действия, которые распространяются по аксону и достигают аксонной терминали. В мембранах этой области нейрона имеются потенциалозависимые (п/з) Nа, К и Са каналы. Nа каналы обеспечивают глубокую деполяризацию мембраны, во время которой активируются Са каналы и в цитоплазму терминали поступают ионы Са. Ионы Са связываются с белком синапсином, который фиксирует везикулы на нитях цитоскелета, вызывают его разрушение и везикулы становятся подвижными.
В мембранах везикул и в пресинаптической мембране имеется система белков, обладающих сродством к Са и регулирующих высвобождение медиаторов. При повышении цитоплазматической концентрации Са повышается активность этих белков, что приводит к слипанию мембран везикул и пресинаптической мембраны. При этом происходит выброс содержимого везикул (иногда и самих везикул) в синаптическую щель. Таким образом, вызванное высвобождение медиатора в любом синапсе является Са-зависимым процессом.
Активация медиатором постсинаптических рецепторов и трансдукция рецепторного стимула
Рецепторы это белковые молекулы, которые распознают свою сигнальную молекулу (медиатор), связываются с медиатором, изменяют свою пространственную структуру и запускают изменяющие функции клетки процессы трансдукции.
7
Постсинаптические рецепторы медиаторов делят на 2 группы: метаботропные и ионотропные рецепторы.
К числу метаботропных рецепторов относят рецепторы ацетилхолина (М-ХР), норадреналина (α- и β-АР), серотонина (все 5-ГТ Р, кроме третьего типа), дофамина (ДА Р), гистамина (Н Р), энкефалинов (μ-, δ- и κ-опиатные Р) и др.
Метаботропный рецептор представляет белок 7 раз пронизывающий постсинаптическую мембрану (рецептор с 7 трансмембранными доменами). Каждый трансмембранный домен содержит 25-35 аминокислотных остатков и все домены соединены тремя внутри- и тремя внеклеточными петлями. Внутриклеточные II и III петли тесно связаны с регуляторными ГТФ-зависимыми G-белками. После связывания медиатора внеклеточные участки трансмембранных доменов сближаются, а внутриклеточные – расходятся и движение внутриклеточных II и III петель вызывает диссоциацию G-белка на α- и βɣ-субъединицы.
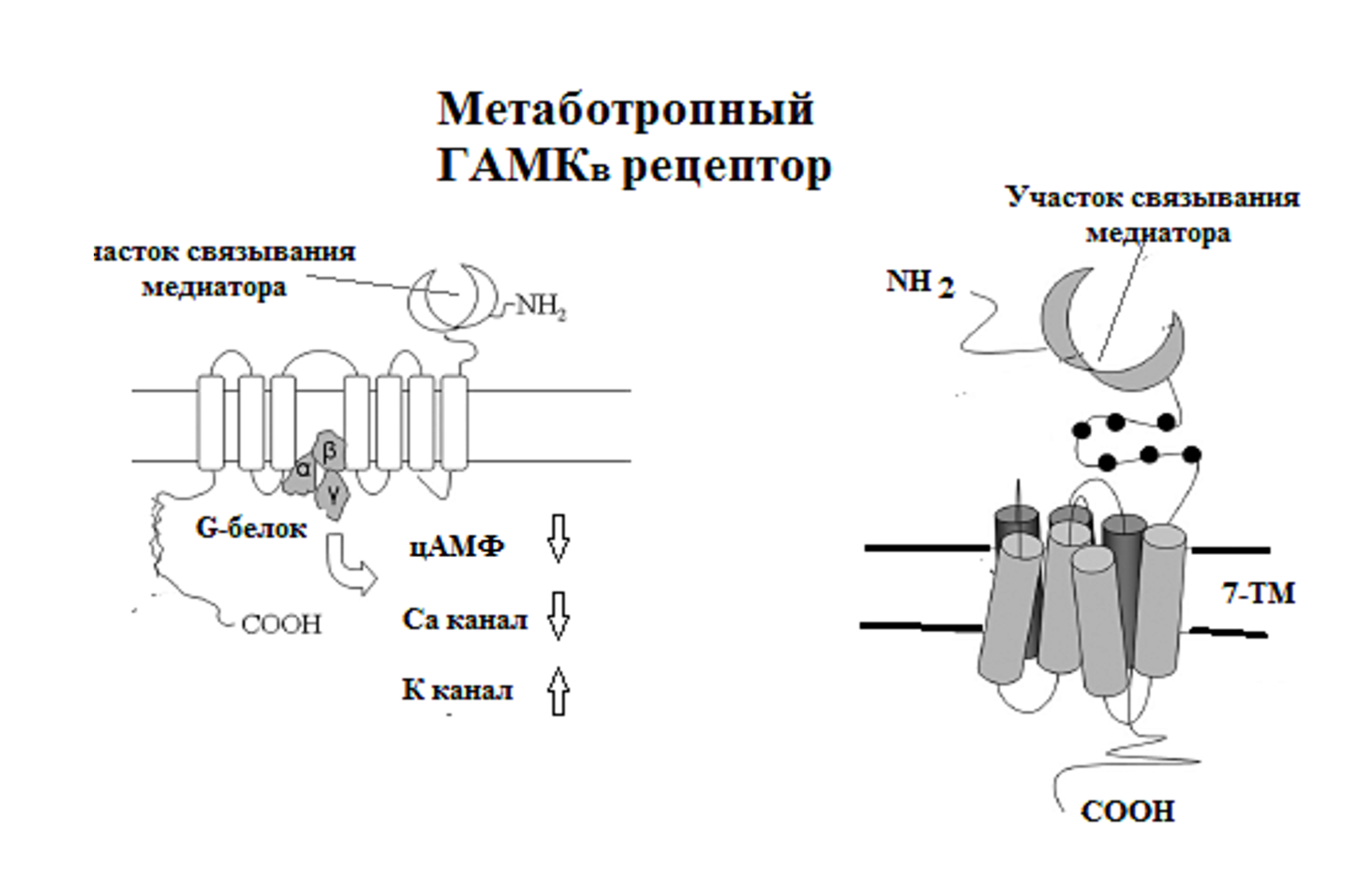
α-субъединица G-белков активирует внутримембранные ферменты: аденилатциклазу или фосфолипазу С, а βɣ-субъединицы взаимодействуют с ионными каналами. Образовавшиеся при активации внутримембранных ферментов вторичные посредники повышают активность протеинкиназ. Действительно, цАМФ активирует протеинкиназу А, Са2+ и ДАГ активируют протеинкиназу С, а Са2+ через посредство кальмодулина активируют Са/кальмодулин-зависимую протеинкиназу. В свою очередь, протеинкиназы фосфорилируют и меняют активность ферментов, транспортеров, ионных каналов и меняют обменные процессы в иннервируемой клетке. βɣ-субъединицы взаимодействуют с К каналами и облегчают их открытие, а также с Са каналами и угнетают их активность.
К
числу ионотропных рецепторов относят
рецепторы ацетилхолина (Н-ХР), глутамата
(АМРА-, НМДА- и КА-Р), серотонина (5-ГТ3-Р),
АТФ (Р2х-Р), ГАМК (ГАМКА-Р) и глицина.
Ионотропные рецепторы представляют
собой сложные, состоящие из нескольких
субъединиц белковые молекулы, пронизывающие
клеточную мембрану. После
связывания медиаторов с рецепторной
частью белка происходит конформационное
изменение структуры всей молекулы
и повышается проводимость ионного
канала рецептора, в результате чего
начинается трансмембранное движение
одного или нескольких ионов по градиенту
концентраций.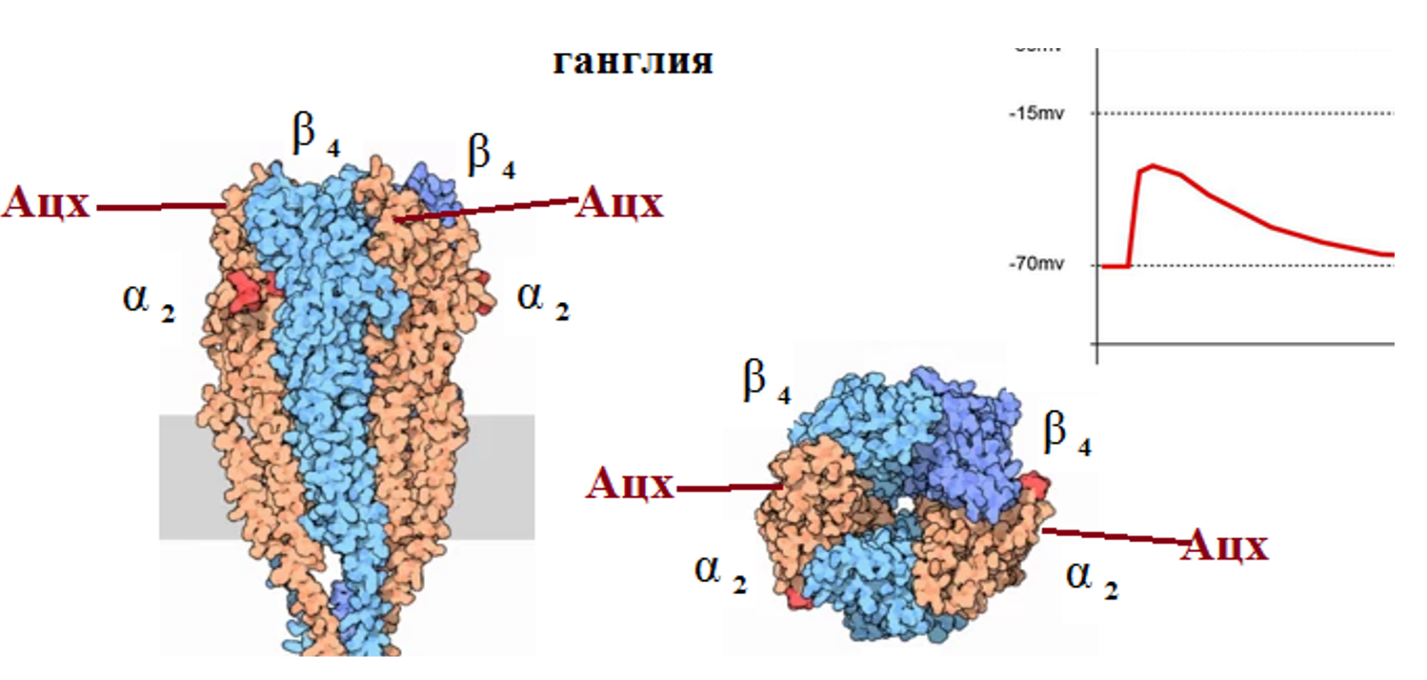
Н-ХР симпатических ганглиев состоит из 5 белковых субъединиц: две α2 и три β4 субъединицы. Две молекулы Ацх связываются с α субъединицами и это вызывает изменение конформации рецептора – происходит вращение всей конструкции против часовой стрелки. В результате увеличивается диаметр центральной ионпроводящей поры и происходит движение ионов Nа и К по градиенту концентраций. При этом возникает входящий трансмембранный ток, который смещает мембранный потенциал в сторону деполяризации и генерируется ВПСП. Если ВПСП достигает критического уровня деполяризации, возникает один или несколько потенциалов действия, которые возбуждают нейрон, вызывают сокращение волокон скелетных мышц и т. п.
9