

Олигосахариды: определение, номенклатура, примеры растительных олигосахаридов. Методы изучения строения олигосахаридов.
Олигосахариды — углеводы с 2-10 моносахаридными остатками.
Номенклатура: строгая громоздка, по типу О-замещенных производных моносахаридов, исходя из названия восстанавливающего звена с указанием всех имеющихся заместителей.
Г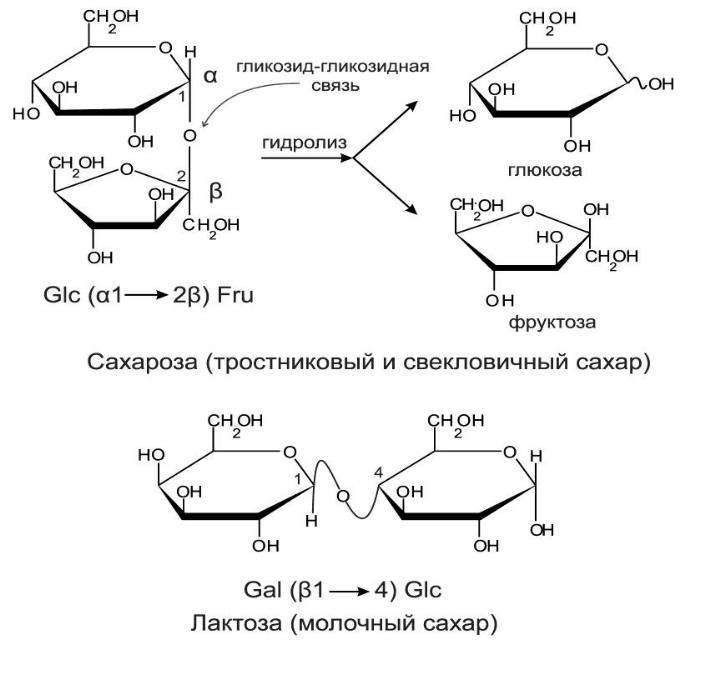
 идроксильная
группа при С1 — повышенная реакционная
способность. При взаимодействии с
электрон-донорами (спирты, амины, тиолы)
они первыми вступают в реакцию конденсации
с образованием воды и аддуктов с общим
названием гликозиды
(углевод
+ другая ФГ). Присоединение спиртов к
аномерному С1 по сути реакция от
полуацеталя до ацеталя, получаемая
связь — гликозидная. При взаимодействии
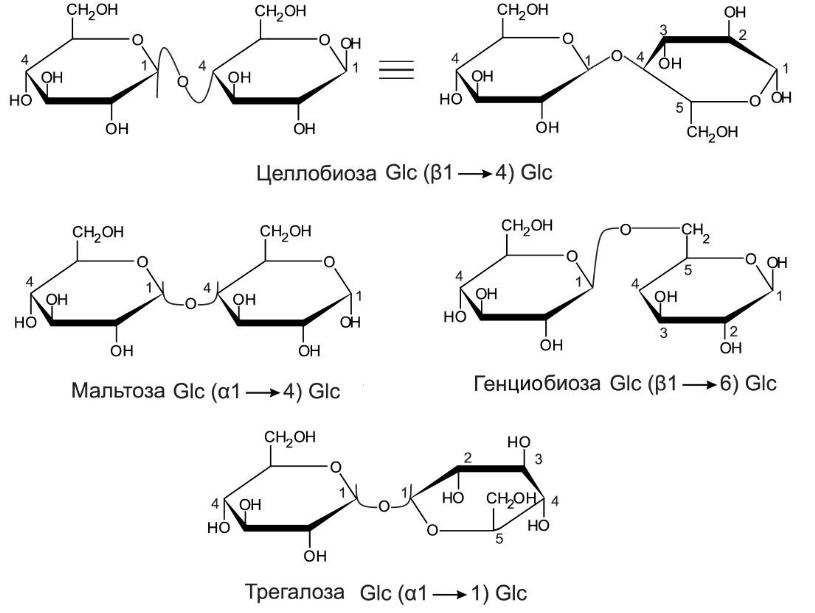
2 моносахаридов = дисахарид с О-гликозидной
связью. Названия производных читают с
мономера, у которого С1 в гликозидной
связи. Если в дисахариде есть доступный
для окисления (свободный) аномерный
атом углерода, он (как и мономер) проявляет
слабые восстанавливающие свойства,
такие сахара — восстанавливающие (при
связи 1-1 невосстанавливающие, как
трегалоза). Трегалоза — источник
"быстрой" энергии для полета
насекомых, основной компонент гемолимфы.
идроксильная
группа при С1 — повышенная реакционная
способность. При взаимодействии с
электрон-донорами (спирты, амины, тиолы)
они первыми вступают в реакцию конденсации
с образованием воды и аддуктов с общим
названием гликозиды
(углевод
+ другая ФГ). Присоединение спиртов к
аномерному С1 по сути реакция от
полуацеталя до ацеталя, получаемая
связь — гликозидная. При взаимодействии
2 моносахаридов = дисахарид с О-гликозидной
связью. Названия производных читают с
мономера, у которого С1 в гликозидной
связи. Если в дисахариде есть доступный
для окисления (свободный) аномерный
атом углерода, он (как и мономер) проявляет
слабые восстанавливающие свойства,
такие сахара — восстанавливающие (при
связи 1-1 невосстанавливающие, как
трегалоза). Трегалоза — источник
"быстрой" энергии для полета
насекомых, основной компонент гемолимфы.
Свойство сахаров восстанавливать металлы — основа их определения; с помощью реактива Бенедикта (щелочной раствор CuSO4) раньше выявляли наличие глюкозы в крови и моче при диагностике диабета. Реактив + моча — нагрев — изменение окраски (зеленая без осадка — проба отрицательная).
Анализ структуры: все разное, всего много, все сложно. В целом:
— чистый сахарид;
— молекулярка;
— определение мономеров;
— определение числа разветвлений;
— сборка полимеров.
Методы: хроматография, химия, ферменты. Масс-спектрометрия и ЯМР-спектрометрия.
Есть химический синтез олигосахаридов для подтверждения их строения. Методы: активация ФГ для конденсации и образования гликозидных связей; защита групп для направленности синтеза; конденсация без мутаротации для нужных аномеров. Синтетические олигосахариды помогают иммунотерапии бактериальных патогенов (выработка высокоаффинных антител к бактериям).
Полисахариды: определение, номенклатура, растительные полисахариды и полисахариды животного происхождения, биологическая роль.
Гликаны — полисахариды и олигосахариды с О-гликозидными связями. Делятся на гомо- и гетерополимеры (с одинаковыми или разными мономерами). У гликанов, в отличие от белков, нет точной молекулярной массы. Сборка гликанов идет вне матрицы, программа синтеза заключена в ферменте-катализаторе, стоп-команда не известна.
Гомогликаны. Глюкоза: крахмал (растительный) и гликоген (животный).
Крахмал: все клетки, больше в клубнях и семенах. Состоит из двух типов полимеров: 1) длинные неразветвленные цепи амилозы (1-4); 2) разветвленные цепи амилопектина (1-4 и через 24-30 остатков ветвление по 1-6).
Гликоген: в целом такой же, но разветвления каждые 8-12 остатков. Запасы гликогена в печени в виде нерастворимых гранул, с ними связан комплекс ферментов для гидролиза и синтеза гликогена. Каждая линейная цепь гликогена заканчивается разветвлением, поэтому у молекулы только один восстанавливающий конец.
Почему клетке удобнее хранить глюкозу в полимерах? Гликоген в гепатоците содержит эквивалентную мономерную глюкозу с С=0,4 М; при такой концентрации растворенной глюкозы неизбежно осмотическое проникновение в клетку кучи воды из м/к пространства (С=5мМ) —> разрыв и лизис. А у нерастворимого гликогена С=0,1мкМ.
Целлюлоза: бета-глюкозы (1-4). Клетки стеблей травы и стволов деревьев, хлопок. В ЖКТ животных а-амилаза гидролизует а1-4 связи, а бета нет, целлюлоза не усваивается. Куча углеводов нам недоступна. Трихонимфа зато декретирует целлюлазу, она есть у жвачных животных в рубке. Целлюлоза линейная, без разветвлений, конформация кресла, каждый следующий мономер повернут вокруг гликозидной связи на 180 гр. => цепи плотно упакованы водородными связями, разлагается сложно.
Хитин: N-ацетилглюкозамин, б(1-4) связи. Длинные волокна, не перевариваются животными. Отличается от целлюлозы только ацетилированным производным аминогруппы по С-2 положению вместо гидроксила. Хитин в основе панцирей членистоногих, чуть ли не второй полисахарид после целлюлозы.