

Эффекты блуждающего нерва
Около двадцати лет назад общепризнанным было то, что блуждающий нерв практически не оказывает влияние на работу желудочков сердца у млекопитающих [2]. Регуляция сократимости миокарда, осуществляемая нервной системой, очень трудно поддается исследованию, так как кроме нейрогенных на нее влияет множество других факторов. Для строгого контроля за этими дополнительными факторами (ЧСС, преднагрузка и постнагрузка, насосная функция предсердий и перфузионное давление в коронарных сосудах) в лаборатории авторов данной главы были разработаны три различных экспериментальных препарата на собаках [55]. Нанесение серии стимулов для раздраже-ния вагуса вызывает уменьшение сократимости желудочков, величина которого обратно пропорциональна частоте стимуляции блуждающего нерва. При максимальной стимуляции вагуса сократимость миокарда желудочков снижается на 15—25%.
Эти результаты были подтверждены на множестве разнообразных препаратов на собаках и на других млекопитающих, включая исследования на людях. Стимуляция вагуса вызывает сдвиг рабочих диаграмм желудочков вправо, существенно уменьшает максимальную скорость повышения внутрижелудочкового давления и снижает силу сокращения миокарда желудочков [56].
Много десятилетий назад Wiggers [57] показал, что предсердия очень чувствительны к влиянию блуждающего нерва и его результаты были полностью подтверждены [58, 59]. Парасимпатическая иннервация предсердий значительно более обильна, чем иннервация желудочков, кроме того, здесь существенно выше содержание холинэстеразы [4, 5]. Поэтому расстояние, на которое вынужден диффундировать Ацх, значительно короче в предсердиях, чем в желудочках, и реакция исчезает быстрее, вследствие более высокого содержания холинэстеразы. Это частично объясняет, почему реакция предсердий на стимуляцию блуждающего нерва описывается меньшими постоянными времени, чем реакция желудочков.
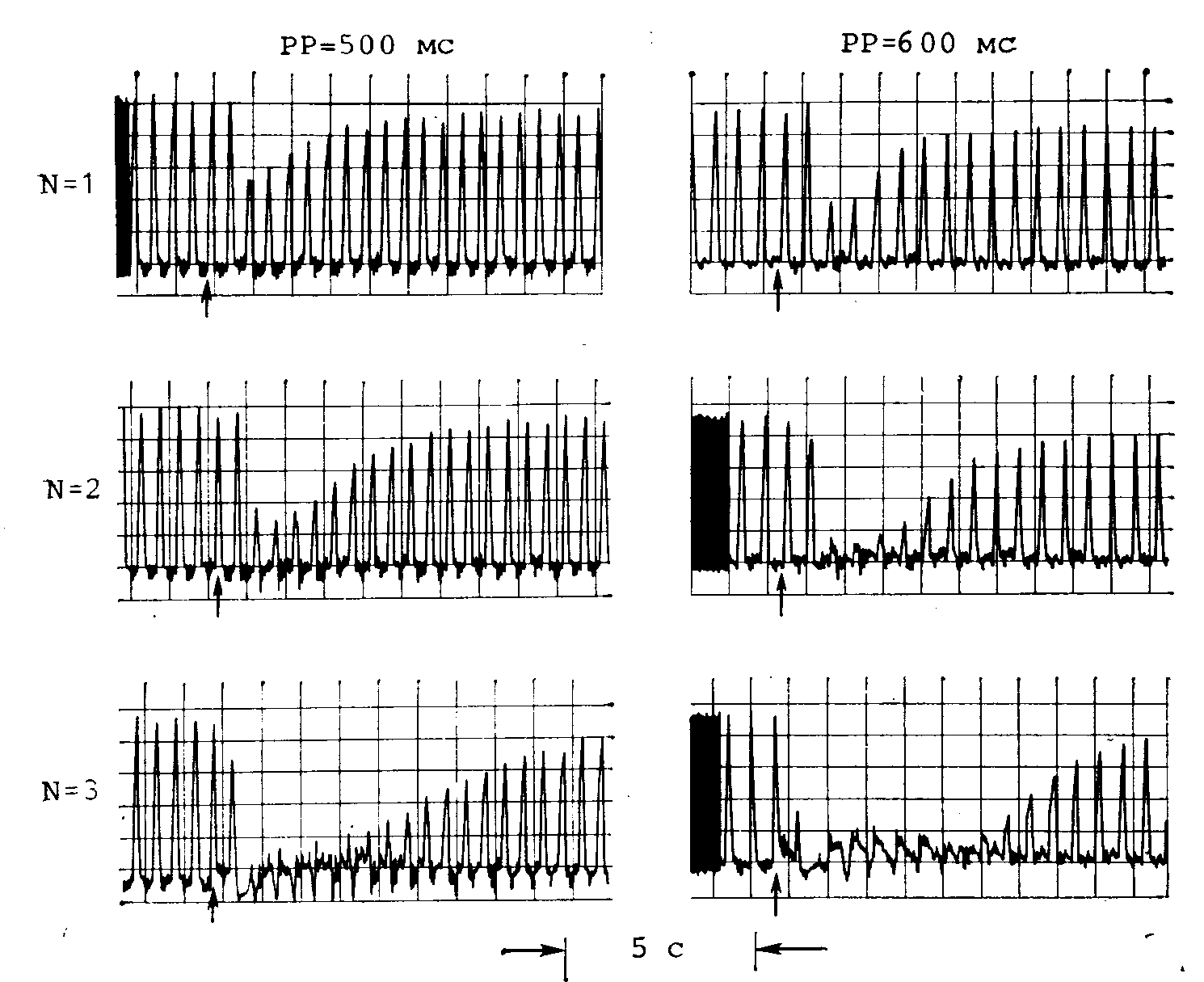
Рис. 22.15. Изменения внутрисердечного давления, регистрируемого предсердным балонным катетером, в ответ на стимуляцию блуждающего нерва пачкой, состоящей из 1, 2 или 3 стимулов (N).
Стимуляцию осуществляли в моменты времени, отмеченные стрелками. Продолжительность сердечного цикла (РР) равнялась 500 и 600 мс. (Заимствовано из работы [60].)
В ранних исследованиях реакций предсердий млекопитающих на повышение активности вагуса использовали длинные серии импульсов и измеряли установившиеся реакции. Недавно провели изучение кратковременных инотропных реакций предсердия собаки на однократную стимуляцию блуждающего нерва или стимуляцию короткой пачкой стимулов [60]. На рис. 22.15 показаны типичные результаты, полученные при нанесении одиночной пачки стимулов, состоящей из одного, двух или трех стимулов, для раздражения вагуса на фоне искусственного ритма сердца двух разных частот. Обратите внимание, что раздражение вагуса одиночным стимулом вызывает уменьшение амплитуды сокращения предсердия на 62% при продолжительности сердечного цикла 600 мс. Пачка стимулов из двух или трех импульсов полностью угнетает сокращение предсердия на 2—8 с. Отрицательная инотропная реакция была меньше при укорочении сердечного цикла до 500 мс.
Для количественного описания инотропных реакций использовали кривые эффекта блуждающего нерва. На рис. 22.16 показаны такие кривые для изменения продолжительности сердечного цикла и сократимости предсердий. Развитие инотропной реакции во времени аналогично временному ходу второго пика хронотропной реакции. Это позволяет предположить, что физиологические механизмы этих двух реакций могут иметь некоторую общую стадию.
Реакция миокарда предсердий на стимуляцию блуждающего нерва зависит также от фазы сердечного цикла, в течение которой подается пачка стимулов [60], точно так же, как это установлено для СП и ПЖ узлов. Вспышка активности вагуса, воздействующая на сердце в течение одной фазы сердечного цикла, может полностью подавить следующее сокращение предсердий, в то время как та же вспышка активности, возникшая в течение другой фазы, может вызвать незначительный эффект. Такое влияние момента возникновения вспышки активности вагуса способно изменять вклад предсердий в наполнение желудочков.
Инотропные реакции предсердий на стимуляцию блуждающего нерва имели большую величину при постоянном ритме сердца, чем когда ритм сердца мог изменяться [60]. Это отличие невозможно объяснить изменением преднагрузки. Реальное объяснение может основываться на свойствах динамической зависимости силы сокращения от ЧСС, которое, вероятно, значительно отличается от обычных характеристик этой зависимости для установившихся условий.
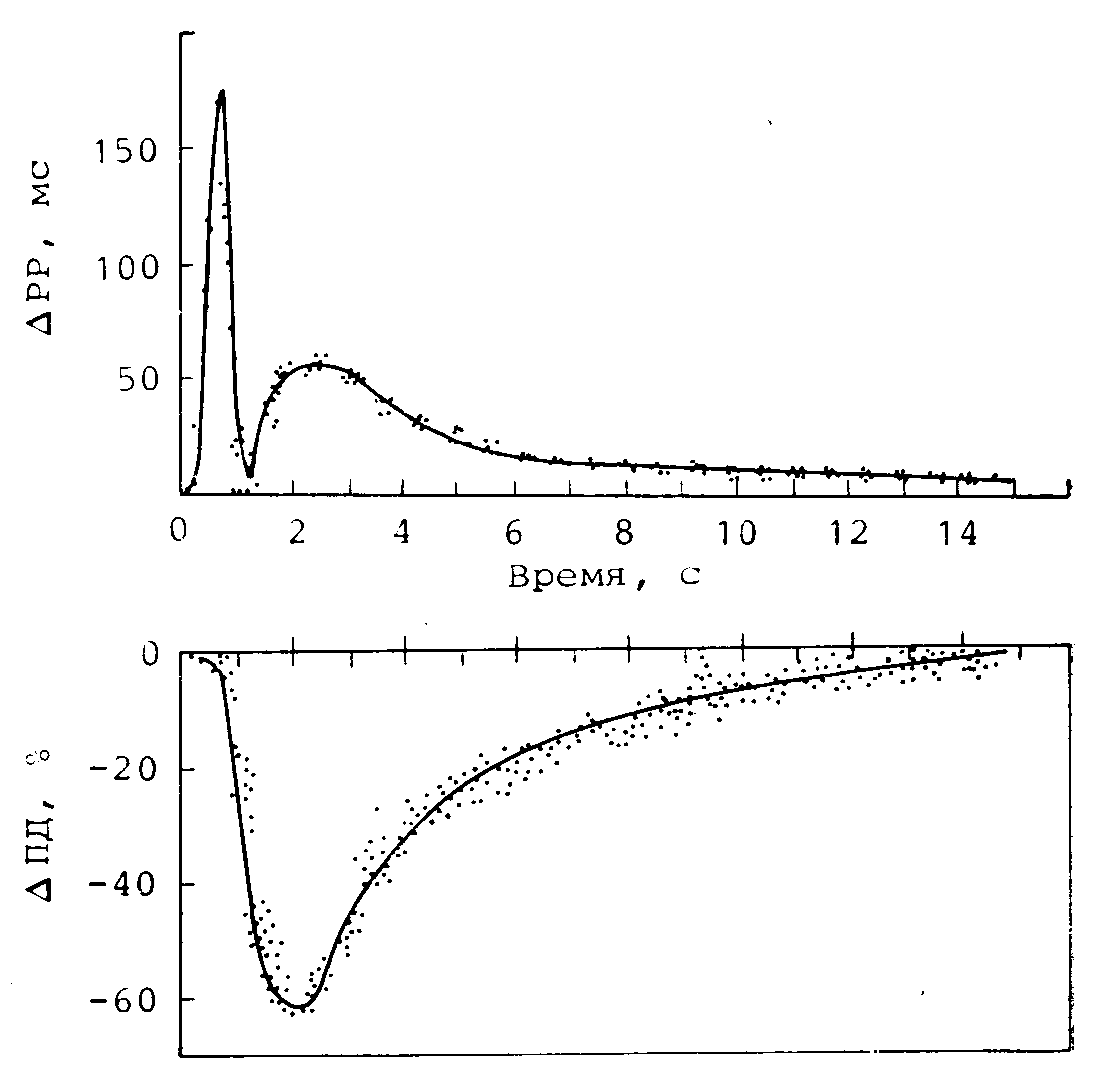
Рис. 22.16. Кривые эффекта стимуляции блуждающего нерва на продолжительность сердечного цикла (РР) при естественном ритме и на давление, измеряемое внутрипредсердным балонным катетером (АР) при искусственном ритме.
Данные получены на одной и той же собаке при нанесении пачек стимулов блуждающего нерва с различными интервалами Р—St (Заимствовано из работы 160].)
Если стимулировать блуждающий нерв в течение нескольких минут с постоянной частотой и интенсивностью [61], длительность сердечного цикла сначала возрастает до некоторой максимальной величины, в то время как сила сокращения снижается до некоторого минимума (рис. 22.17). Однако постепенно обе величины восстанавливаются до контрольных значений. Уменьшение длительности сердечного цикла до контрольного уровня наблюдается приблизительно через 15—20 с, в то время как восстановление силы сокращений происходит за 1—2 мин. Однако реакция ПЖ проведения во время непрерывной стимуляции вагуса не исчезает [62]. По-видимому, исчезновение различных типов реакций сердца на фоне постоянной стимуляции блуждающего нерва определяется различными механизмами. Имеющиеся в настоящее время данные позволяют предположить, что значительную роль при этом играет уменьшение чувствительности мускариновых рецепторов к Ацх [63], т. е. рецепторы постепенно теряют свою способность связывать агонист, хотя нельзя полностью исключить и постепенного уменьшения освобождения Ацх из нервных окончаний во время непрерывной стимуляции.
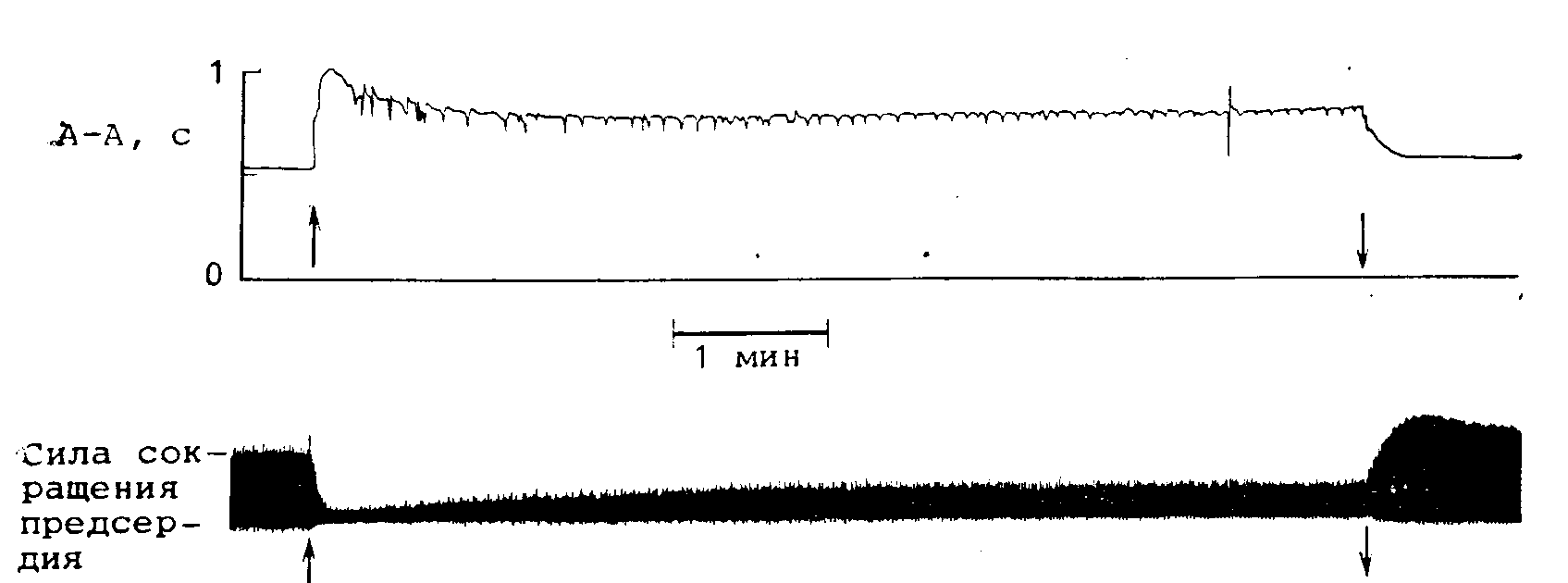
Рис. 22.17. Изменения продолжительности сердечного цикла (вверху) и силы сокращения предсердия (внизу) в ответ на непрерывную стимуляцию блуждающего нерва (между стрелками).
Обратите внимание, что несмотря на непрерывную стимуляцию вагуса хронотропная и ннотропная реакции постепенно возвращаются к контрольному уровню после достижения некоторого пикового значения. (Заимствовано из работы [61].)
РЕФЛЕКТОРНАЯ РЕГУЛЯЦИЯ СЕРДЕЧНОЙ ДЕЯТЕЛЬНОСТИ
Рефлекторная регуляция функций сердца изучалась очень интенсивно, и недавно было опубликовано несколько исчерпывающих обзоров по этому вопросу [64—68]. Здесь приводится лишь краткое описание трех наиболее важных рефлексов; барорецепторного рефлекса, рефлекса Бейнбриджа и хеморецептор:ного рефлекса.
Барорецепторный рефлекс
Барорецепторы представляют собой рецепторы, воспринимающие механическое растяжение стенки артерий, и расположены в каротидных синусах и дуге аорты [65, 68]. Афферентные импульсы от рецепторов каротидных синусов поступают в головной мозг по нервам каротидных синусов, которые являются ветвями языкоглоточных (IX пара черепных нервов). Импульсы от барорецепторов аорты поступают в мозг по аортальным нервам, которые являются ветвями блуждающих нервов (X пара черепных нервов). Эфферентное плечо барорецепторного рефлекса образуется симпатическими и парасимпатическими волокнами.
Частота импульсации барорецепторов стенок артерий увеличивается при повышении среднего артериального давления в области каротидных синусов и дуги аорты. Увеличение частоты импульсации в нервах каротидных синусов и аортальных нервах вызывает уменьшение нервной активности в эфферентных симпатических волокнах и увеличение активности в эфферентных парасимпатических волокнах. Снижение симпатической активности уменьшает вазомоторный тонус в резистивных и емкостных сосудах всего тела, способствует понижению ЧСС, увеличивает время ПЖ проведения и уменьшает сократимость предсердий и желудочков. Повышение активности блуждающего нерва вызывает те же эффекты, что и снижение симпатической активности.
Противоположные изменения эфферентной симпатической в парасимпатической активности в ответ на изменение артериального давления наблюдаются только тогда, когда артериальное давление находится вблизи нормального диапазона давлений. Если артериальное давление резко снижается до аномально низкого уровня, тонус блуждающего нерва практически исчезает. В этом случае градация рефлекторной регуляции осуществляется исключительно за счет изменений эфферентной симпатической активности. И наоборот, если артериальное давление резко повышается до аномально высокого уровня, симпатический тонус полностью угнетается, а градация рефлекторной регуляции осуществляется только за счет изменений эфферентной активности вагуса.