

4.3. Залежність між осмотичними і тургорним тиском, та водним потенціалом
При поглинанні клітиною води цитоплазма притискується до клітинної стінки, виникає протитиск клітинної оболонки на вміст клітини, створюється гідростатичний тиск. Активність води під впливом тиску зростає, тому гідростатичний потенціал має позитивне значення.
При повному насиченні водою (максимальний тургорний тиск) позитивний потенціал тиску врівноважує негативний від’ємний потенціал, тобто водний потенціал клітини дорівнює нулю. При такому стані поглинання води не відбувається. Якщо ж осмотичний потенціал перевищує потенціал тиску, то водний потенціал буде мати негативне значення, і клітина здатна поглинати воду.
Надходження води визначається різницею між осмотичним (Р) і тургорним (Т) тиском. Цю величину називають всисною силою (S):
S = Р – Т.
Процес поглинання води клітиною триває до повного насичення, доки не буде досягнуто найбільше розтягування клітинної оболонки. Клітина при цьому матиме максимально можливий об’єм, концентрація клітинного соку стане мінімальною, а тургорний тиск – максимальним. Якщо таку клітину помістити у концентрований розчин, вона втрачатиме воду. Об’єм вакуолі зменшиться, зменшиться і тиск цитоплазми на клітинну оболонку, внаслідок чого знизиться тургорний тиск. При досягненні клітиною мінімального об’єму і подальшій втраті води протоплазма почне відставати від оболонки. Процес втрати клітиною води буде тривати до зрівноваження концентрацій клітинного соку і зовнішнього розчину. Явище відставання протоплазми від клітинної оболонки називається плазмолізом, а клітина – плазмолізованою (рис. 17).
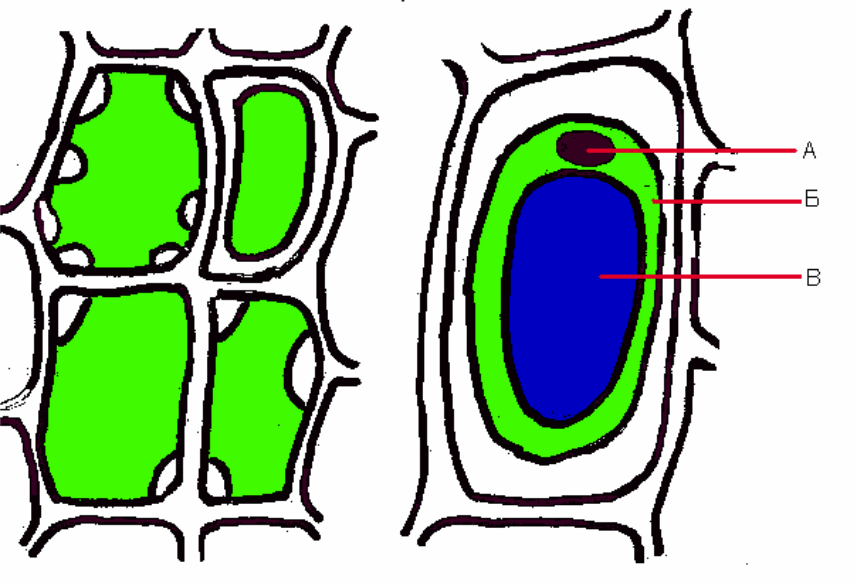
1 2
Рис. 17. Плазмоліз рослинної клітини: 1 – послідовні стадії плазмолізу у клітинах моху; 2 – опукла форма плазмолізу (ковпачковий плазмоліз клітини епідермісу луски цибулі; а – ядро; б – цитоплазма; у – вакуоля.
Якщо плазмоліз незначний і нетривалий, то при наявності вологи клітина може відновити тургор. При нестачі вологи у повітряному середовищі і втраті клітиною води між протопластом і клітинною оболонкою не утворюється вільний простір, але протопласт, зменшуючись у об’ємі, тягне за собою клітинну оболонку. Поверхня клітини стає хвилястою. Такий стан клітини називається циторизом. У прив’ялих рослин тургорний тиск клітин стає меншим нуля, тобто від’ємною величиною. Це пояснюється тим, що при циторизі сили пружності клітинної оболонки не стискують протопласт, а розтягують. Сума сил циторизу й осмотичного тиску визначає величину всисної сили зів’ялої клітини:
S = Р + Т.
Зміни осмотичних показників у клітині при переході від стану плазмолізу до повного насичення водою показано на рисунку 18. Ліва частина рисунка відповідає стану повного в’янення, або плазмолізу, коли цитоплазма має найменший об’єм. При цьому осмотичний тиск (Р) – максимальний, тургорний (Т) – відсутній, а величина всисної сили (S) досягає найбільшого значення.

Рис. 18. Залежність між всисною силою (S), тургорним (Т) і осмотичним (Р) тиском.
Поглинаючи воду, така клітина буде збільшуватися в об’ємі, концентрація клітинного соку і осмотичний тиск зменшаться, з’явиться і почне зростати тургор. У правій частині рисунка при повному насиченні клітини водою тургорний (Т) і осмотичний тиск (Р) зрівнюються, а всисна сила (S) дорівнюватиме нулю. Таким чином, рослинна клітина є саморегульованим осмотичним механізмом.
Осмотичні властивості клітини не постійні й залежать від видових особливостей рослин, типу тканин, багатьох внутрішніх і зовнішніх факторів. При вирощуванні рослин в умовах з низькою вологістю ґрунту і повітря осмотичний тиск клітинного соку вищий, ніж у тих, що вирощуються при сприятливих умовах водозабезпечення. У рослин різних екологічних груп осмотичний тиск вищий у ксерофітів, а у гігрофітів – низький.