

2.2. Моделирование численности взаимодействующих популяций.
Предыдущий частный пример - ограничение роста численности абиотическими факторами. Более интересные и важные для эволюции и экологии ситуации возникают при взаимодействии популяций разных видов (перекрывание экологических ниш) или при изменениях внешних условий. В подобных ситуациях по численности Nt возникают так называемые популяционные волны или волны жизни (по С.С Четверикову).
Классификация популяционных волн:
Периодические колебания численности популяции (например, сезонные).
Непериодические или периодические колебания численности популяций, вызванные, например, взаимодействием популяций жертвы и хищника.
Вспышки численности (когда популяция попадает в благоприятные условия).
Резкие сокращения численности популяции (эпифитотии, катастрофы).
Возможны несколько типов взаимодействия двух популяций разных видов: (-, -) – конкуренция, при которых ухудшаются условия жизни для обеих популяций; (+,+) – симбиоз; (+,-) – хищник-жертва и т.д.
Рассмотрим взаимоотношение типа «хищник-жертва». Проследим, как изменится численность популяций хищника и жертвы, если поместить их в ограниченный объём с достаточным количеством пищи для жертв.
Пусть х – численность жертвы; у – численность хищника. Тогда для моделирования можно воспользоваться уравнениями Лотки - Вольтерра:
![]()
Здесь ху – характеризует частоту встречи жертвы и хищника в их ограниченном ареале совместного обитания;
r – скорость естественного увеличения популяции жертв (без учета влияния хищников);
Kmax - предел увеличения численности жертв в ограниченном ареале (число хищников обычно гораздо меньше числа жертв);
с – коэффициент успеха охоты;
g – коэффициент рождаемости для хищников (скорость увеличения их численности зависит не только от х, но и у, точнее пропорциональна ху);
f – коэффициент естественной смертности хищников;
Решения этих уравнений – волновые колебания численности хищника и жертвы. Их форма и периодичность зависят от начальных условий (х0, у0), а также от констант с, f, r, g и Kmax.
Возможно несколько вариантов:
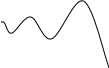
Выход на стабильный уровень (рис. б). Такая ситуация наблюдается, когда хищникам для Nty=const нужно приблизительно столько жертв, сколько их рождается сверх Ntx=const.
Рис. б
x,y

x0
y0

t
Интенсивное поедание жертв, а затем и гибель хищника от голода. Волны идут «вразнос» по амплитудам, пока х не станет равным 0 (рис. в).
Рис. в
x,y
x0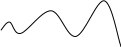
y0

t
Cтабильные волны с постоянными амплитудами (рис. г).
Рис. г
Error: Reference source not found
x,y
Эта математическая модель была подтверждена экспериментами на простейших (Смит, 1976), которые послужили физической моделью для изучения обычных экологических и эволюционных ситуаций в природе. Два вида реснитчатых (популяции хищника и жертвы) были помещены в ограниченный объем жидкости (колбу) с добавлением достаточного количества пищи для жертв. Можно было наблюдать разные ситуации:
Если в колбе не было хищников, то рост численности популяции жертв происходил до Kmax., определяемым объемом жидкости. Форма кривой численности жертв была аналогична рис. а.
При добавлении в колбу популяции хищника последние активно поедали жертв, увеличивая свою численность. Численность жертв при этом сокращалась, пока они не исчезали полностью, что в свою очередь, приводило к гибели популяции хищника от голода (рисунок).
x,y


x0
y0

t
Для того чтобы снизить коэффициент успеха охоты для хищника (с), а значит и коэффициент рождаемости для хищника (g), в данном эксперименте в жидкость добавили целлюлозу, которая увеличивала вязкость раствора. В этом случае возникали волны с нарастающей амплитудой до тех пор, пока все жертвы всё же не были съедены (х=0), что повлекло за собой гибель хищников (рис. в).
Для сокращения скорости естественного увеличения популяции жертв (r) снизили в 2 раза содержание корма для жертв. В этом случае амплитуда увеличения числа жертв стала значительно меньше и, как следствие, не наблюдалось ответного резкого увеличения численности хищника и, как следствие, численность жертв резко не снижалась. Возникали стабильные волны по х и у (рис. г).
Аналогичные модели применимы, например, для объяснения и описания колебаний численностей в системе “хозяин – паразит”.