

7.3.2.3. Этиоляция и деэтиоляция
У цветковых растений в отсутствие условий для фотосинтеза хлорофилл не образуется, не происходит сборка светособирающих комплексов, пластиды имеют агранальную структуру (этиопласты) (см. подразд. 3.2.3). Растения, выросшие в темноте, становятся белыми или желтоватыми, междоузлия удлинены, а листовые пластинки часто превращаются в чешуи. Процесс «обесцвечивания» растений при недостатке света называют этиоляцией. Этиоляция — адекватная реакция на недостаток света: фотосинтетическая функция сведена к минимуму, рост происходит за счет растяжения (форма роста, требующая минимум пластических веществ) (рис. 7.29).
* PAS-домен назван по первым буквам названий генов, в которых он был впервые обнаружен: PER, ARNT, SIM.
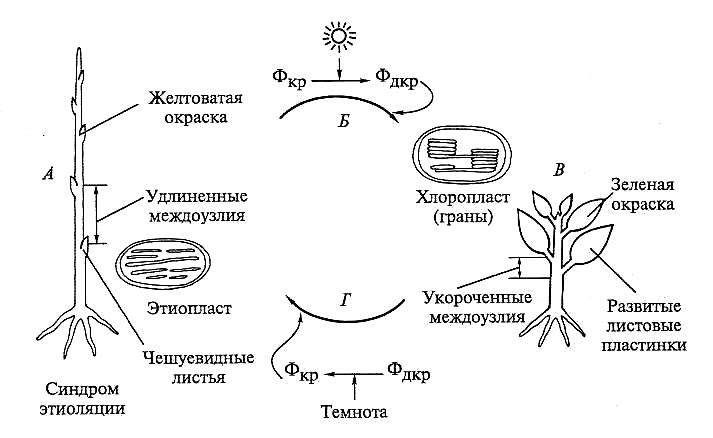
Рис. 7.29. Синдром этиоляции:
А — этиолированное растение; Б — процесс деэтиоляции, регулируемый светом через фито-
хромную систему; В — деэтиолированное растение; / — этиоляция возникает при недостатке
света; сигнал поступает через фитохромную систему
Растение «экономит» органические вещества, «отказавшись» от роста пластинок листьев.
Отсутствие света при прорастании является сигналом, что растение, вероятно, находится под землей, и проросток формирует апикальную петельку, защищающую меристему от возможных механических воздействий. В отличие от тройного ответа на этилен, гипокотиль при этиоляции интенсивно растет.
В результате усиленного роста растение может оказаться в зоне с благоприятным освещением. Как только оно получит сигнал, что свет пригоден для фотосинтеза, у листьев появляется развитая пластинка (что контролирует фитохром С), рост междоузлий ингибируется, синтезируется хлорофилл и пластиды превращаются в хлоропласты, разгибается апикальная петелька, а рост ги-покотиля тормозится. Цепь этих процессов называют деэтиоляцией. Деэтиоляция характеризуется не только синтезом хлорофилла, но и экспрессией генов белков светособирающего комплекса (хлорофилл a/b-связывающего белка). Благодаря гидрофобным свойствам хлорофилл а/b-связывающего белка тилакоиды, содержащие ССК, собираются в граны.
7.3.2.4. Избегание тени
Ситуация, когда растение попадает на свет, обогащенный ДК-лучами, заметно отличается от полной темноты. С одной стороны, дальний красный свет (как и просто красный) вызывает переход фитохрома А в форму Ф(А)730 (это означает, что растение почувствовало свет). С другой стороны, дальним красным светом невозможно перевести Ф(В)660 в Ф(В)730. Растение получает два противоречащих друг другу сигнала: от фитохрома А — о том, что свет есть, а от фитохрома В — о том, что этот свет не пригоден для фотосинтеза. Ответ зависит от «интегрирования» этих взаимно противоположных сигналов.
В природе интенсивный свет, обогащенный ДК, бывает под пологом растений. В нижних ярусах много дальнего красного, так как листья поглощают красные кванты в процессе фотосинтеза. Много дальнего красного — это сигнал о том, что рядом есть более высокие соседи. Однако есть надежда, что рядом обнаружится свободное светлое «окно», не занятое другими растениями.
Если растение получило положительный сигнал от фитохрома А и отрицательный от фитохрома В, развивается «синдром избегания тени». Для этого синдрома характерны черты как этиоляции, так и нормального развития на свету. Хлорофилл синтезируется, но его содержание меньше. Листовые пластинки формируются, но они не такие крупные, как при полном освещении. Удлинение междоузлий менее интенсивно, чем в темноте, но все же скорость роста выше, чем на прямом солнце. В результате образуются длинные побеги со слабо развитыми механическими тканями. Они легко полегают, при этом меристема побега оказывается на новом месте. Происходит новая «оценка ситуации», если по-прежнему в спектре много дальнего красного и мало красного, снова развивается побег с «синдромом избегания тени».
Полное развитие фотосинтетического аппарата возможно только в том случае, когда на растение падает много красного света (и он не обогащен дальним красным). Тогда и от фитохрома А, и от фитохрома В приходят одинаковые сигналы о том, что свет есть и он пригоден для фотосинтеза.
Образно можно назвать фитохром А «оптимистическим» рецептором, а фитохром В — «критиком», или «пессимистом».
Интересно, что поведение растений в тени заметно отличается у мутантов по фитохрому А и В. Если в растении поврежден фитохром В, оно развивает нормальный «синдром избегания тени» и в конечном итоге выживает. Мутанты по фитохрому А при интенсивном дальнем красном освещении сильно вытягиваются и погибают. Этот пример показывает, что отсутствие «оптимизма» бывает очень опасным, и даже смертельным при определенных внешних обстоятельствах. Растению нужно не только «уметь правильно и критически оценить ситуацию» с освещением (функция Ф(В)), но и «почувствовать положительные тенденции — понять», что свет все-таки есть и «жить стоит» (функция Ф(А)). Кроме того, синдром избегания тени регулируется фитохромами D и E.