

Конечный мозг
КОНЕЧНЫЙ МОЗГ — telencephalon — состоит из двух полушарий большого мозга — hemispherium dextrum et sinistrum, разделенных с дорсальной поверхности глубокой продольной щелью — fissura longitudinalis cerebri.
В каждом полушарии рассматривают плащ, обонятельный мозг, полосатые тела и боковые желудочки мозга. Плащ расположен в полушарии дорсо-латерально от желудочка, обонятельный мозг — вентро-медиально. Границей между плащом и обонятельным мозгом на вентральной поверхности мозга служит базальная пограничная, или обонятельная, борозда — sulcus rhinalis. Полосатое тело лежит н вентральной стенке полушария дорсально от частей обонятельного мозга.
А) Плащ — pallium — состоит из серого и белого мозгового вещества. Серое мозговое вещество — substantia grisea — образует кору большого мозга — соrtex cerebri — на его поверхности. На ней различают извилины мозга — gyrus, разделенные бороздами — sulci – и щелями — fissurae. На латеральной поверхности плаща видны три дугообразные извилины. Они окружают латеральную сильвиеву борозду — sulcus lateralis. Дугообразные извилины четко выражены у собаки, в то время как у других домашних животных каждая дуговая извилина подразделяется бороздами на части (рис. 13). На медиальной поверхности плаща в основном заметны две дугообразные извилины вокруг мозолистого тела, причем ближайшая к нему поясная извилина — gyrus cinguli, s. limbicus — функционально связана с обонятельным мозгом.
Белое мозговое вещество плаща — substantia alba — располагается под корой плаща. Оно со стоит из проводящих путей ассоциативных, комиссуральных и проекционных.
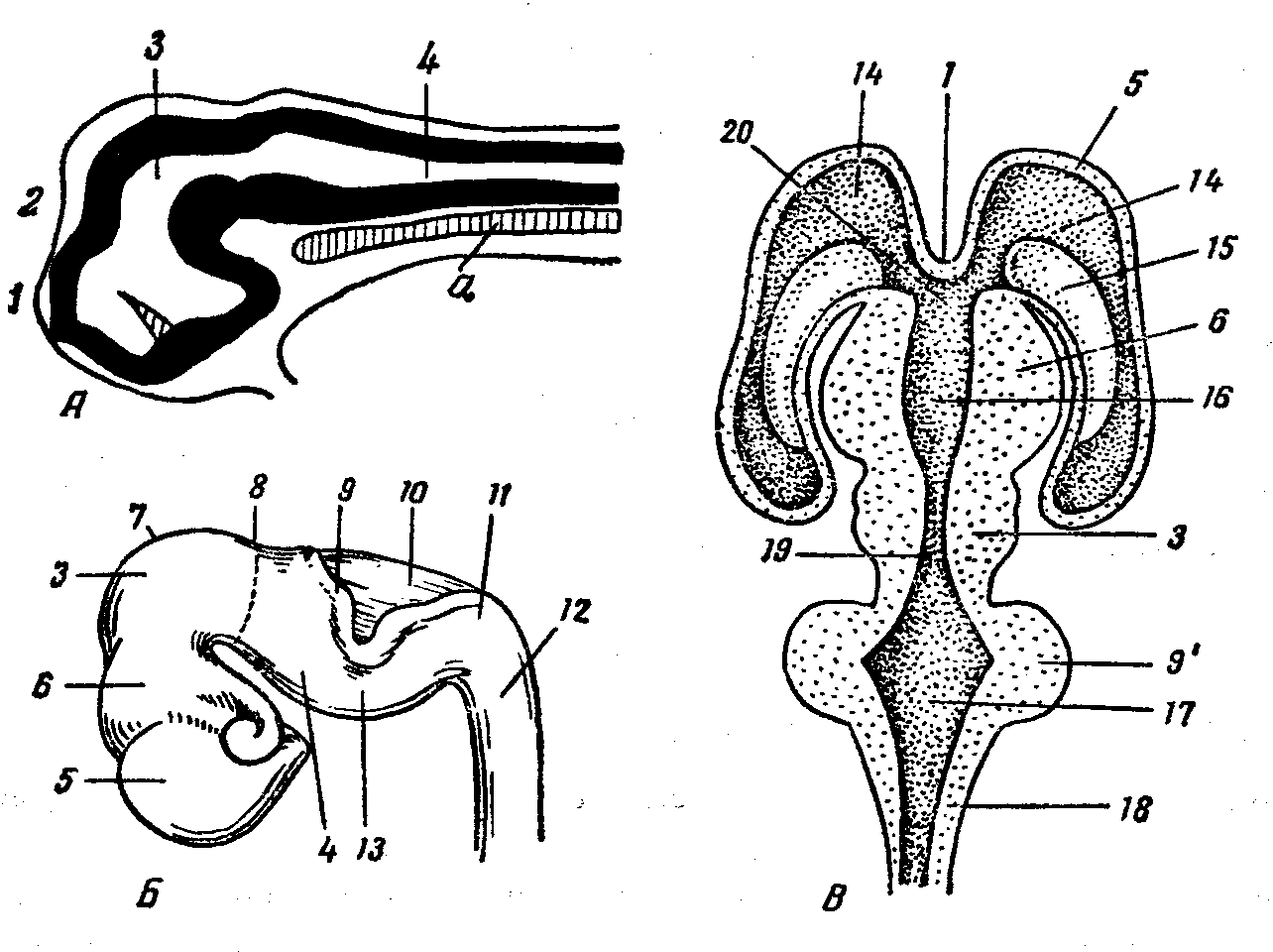
Рис. 11. Развитие головного мозга:
А — трехпузырная и Б — пятипузырная стадии, В — схема желудочков мозга; 1 — концевая пластинка, 2 — передний мозг (прехордальный мозг), З — средний мозг, 4 — ромбовидный мозг, З и 4 — эпихордальный мозг, 5 — конечный мозг, 6 — промежуточный мозг, 7 — теменной бугор, 8 — перешеек, 9 — закладка мозжечка, 9’ — мозжечок, 10 — мозговой парус, 11 — затылочный изгиб, 12 — спинной мозг, 13 – мостовой изгиб, 14 — боковой желудочек, 15 — полосатое тело, 16 — третий мозговой желудочек, 17 — четвертый мозговой желудочек, 18 — продолговатый мозг, 19 — мозговой водопровод, 20 — межжелудочковое отверстие, а — хорда
1) Ассоциативные волокна соединяют отдельные участки коры в пределах каждого полушария. Они разделяются на короткие волокна (между извилинами) и длинные (между долями полушарий).
2) Комиссуральные волокна соединяют участки, принадлежащие разным полушариям. Они формируют мозолистое тело — corpus callosum – самую крупную комиссуру головного мозга (рис. 14, 15). Оно помещается между полушариями в глубине продольной щели. Различают ствол мозолистого тела — truncus corporis callosi — и два конца — передний и задний. Передний конец называется коленом мозолистого тела — genu corporis callosi; он загибается вентрально. Задний конец, или валик, мозолистого тела — splenium corporis callosi — срастается со сводом.
Комиссуральные волокна, выходящие из ствола мозолистого тела, образуют лучистость спайки — radiatio corporis callosi (рис. 15); она формирует дорсальную стенку бокового желудочка мозга и расходится в передний, боковой и задний отделы коры плаща.
3) Проекционные волокна — соединяют кору плаща как с отдельными частями ствола головного мозга, так и со спинным мозгом. Они образуют в полосатом теле внутреннюю капсулу.
Функционально проекционные пути разделяются на эфферентные и афферентные.
Эфферентные проводящие пути (центробежные, двигательные) выносят импульсы из коры полушарий большого мозга в разные отделы ствола мозга: большого, ромбовидного и спинного.
Афферентные проводящие пути (центростремительные, чувствительные) приносят импульсы в кору полушарий из спинного и ромбовидного мозга. Они образуют заднюю часть внутренней капсулы.
В противоположность коре больших полушарий все серое мозговое вещество остальных отделов всей центральной нервной системы объединяется понятием «подкорка». Импульсы со всех частей тела сначала поступают в разные отделы подкорки, включая зрительный бугор, а из последнего уже поступают в кору полушарий. Кора постоянно разлагает, расчленяет возбуждения, возникающие в рецепторных приборах, и в то же время объединяет, синтезирует действие выделенных анализом возбуждений в разнообразные комплексы. Это и представляет собой проявление аналитической и синтетической деятельности коры головного мозга. Более того, высший отдел нервной системы — кора большого мозга — является (у человека) материальным субстратом высшей нервной деятельности: мышления, сознания, памяти, воли, рассудка, т. е. всей психической деятельности, которая представляет собой результат физиологической деятельности коры головного мозга.
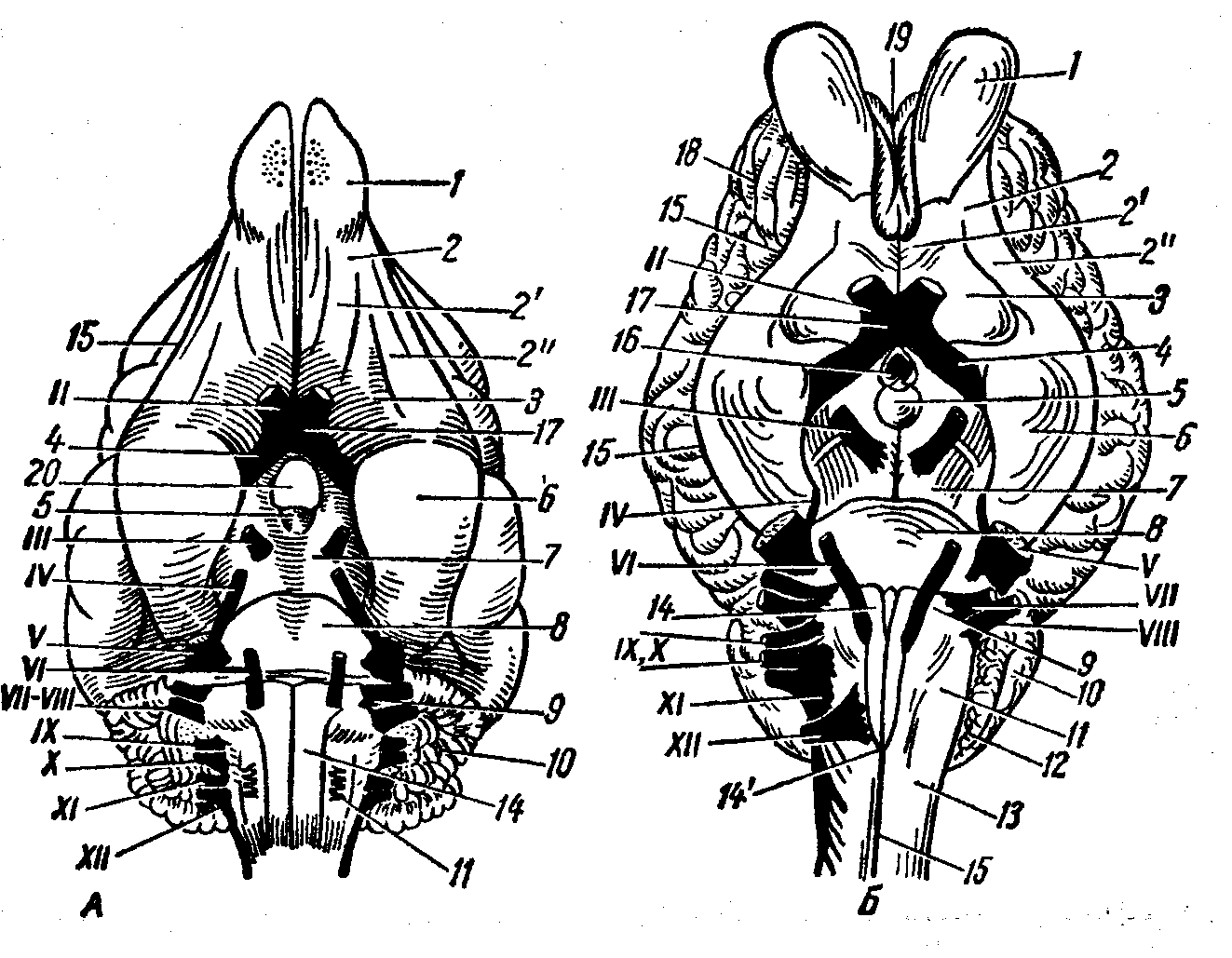
Рис. 12. Головной мозг с базальной поверхности:
А — собаки, Б — лошади; 1 — обонятельная луковица, 2 — общий, 2’ — медиалъный и 2’ — латеральный обонятельные тракты и извилины, З — обонятельный треугольник, 4 — зрительный тракт, 5 — сосцевидное тело, 6 — грушевидная доля, 7 — ножка большого мозга, 8— мозговой мост, 9 — трапециоидное тело, 10 — мозжечок, 11 — продолговатый мозг, 12 — сосудистое сплетение четвертого мозгового желудочка, 13 — спинной мозг, 14 — пирамида, 14’— перекрест пирамид, 15 — базальная борозда, 16 — серый бугор и воронка, 17 — зрительный перекрест, 18 — плащ, 19 — обонятельная медиальная борозда, 20 — гипофиз, II—ХII — черепные нервы.
На плаще рассматривают доли: лобную, височную, теменную, затылочную и обонятельную. Лобная доля — lobus frontalis pallii — лишь у собаки четко отграничивается венечной бороздой — sulcus coronalis. Затылочная доля — lobus оссiрitalis pallii — занимает каудальный отдел плаща, позади плоскости, проведенной через валик мозолистого тела. Теменная доля — lobus parietalis pallii — лежит между лобной и затылочной долями. Височная доля — lobus temporalis pallii — находится приблизительно позади латеральной борозды (сильвиевой) в вентральной половине плаща. Обонятельная доля — lobus olfactorius pallii — образует обонятельный мозг.
Б) Обонятельный мозг — rhinencephalon, s. lobus olfactorius — располагается в вентро-медиальном отделе каждого полушария большого мозга. Отдельные части его видны на базальной и медиальной поверхностях полушарий, а также на дне боковых желудочков мозга (рис. 11, 13, 14, 15).
На базальной поверхности полушарий располагаются обонятельные луковицы, обонятельные тракты и извилины, обонятельные треугольники и грушевидные доли. На медиальных поверхностях полушарий видны околообонятельное поле, извилины гиппокампа, поясные извилины и поверхность разреза назальной спайки, а на дне боковых желудочков мозга— хвостатые ядра, гиппокамп и свод. Поясная извилина, гиппокамп, свод и миндалевидное ядро составляют pars limbica rhinencephali — лимбическую систему.
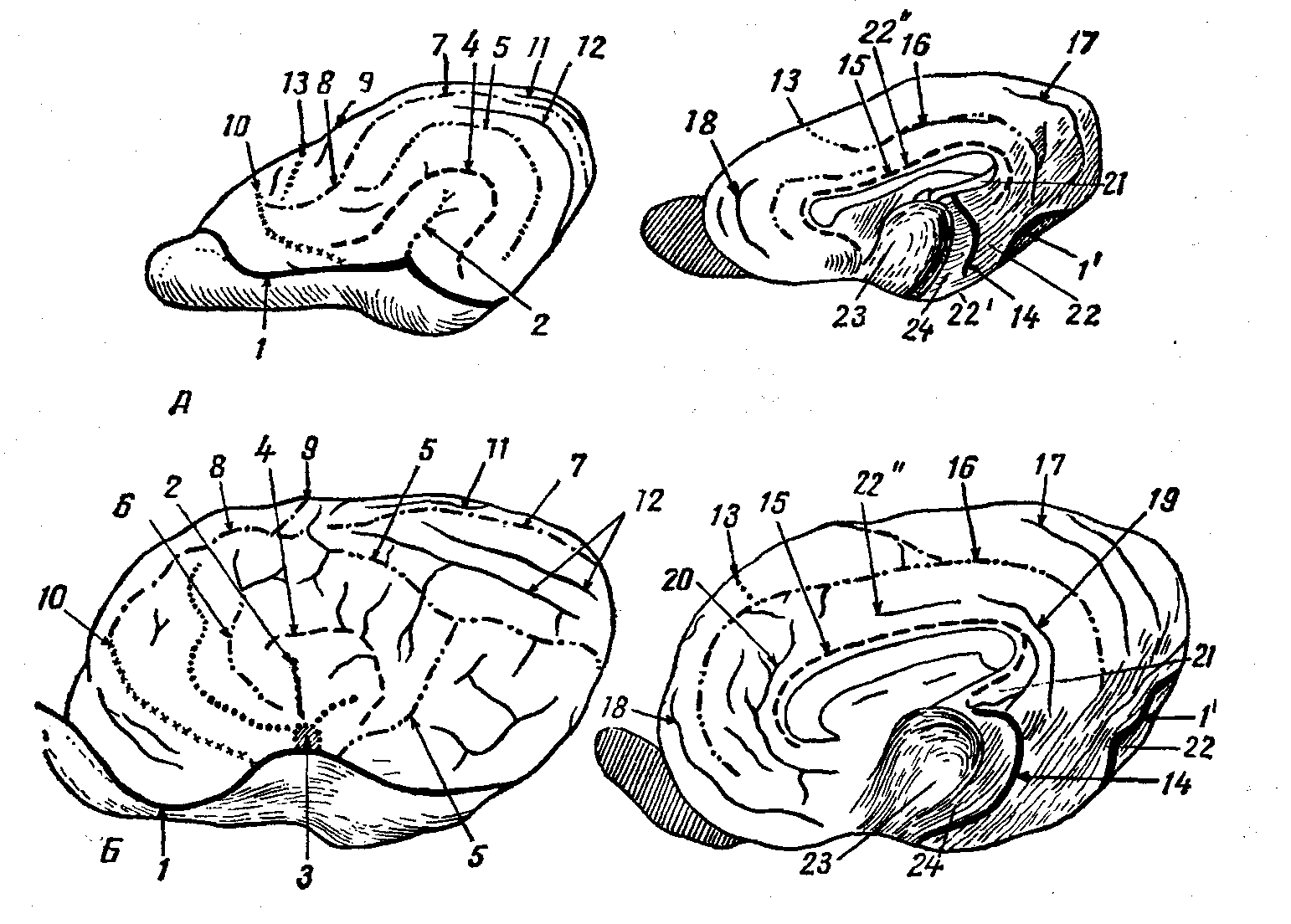
Рис. 13. Схема борозд и извилин плаща:
А — собаки, Б — лошади; (Слева — латералькая поверхность, справа — медиальная.) 1 — обонятельная борозда, 1’ — ее продолжение как каудальная часть, 2 — сильвиева борозда, З — островок Рейля (крючок), 4 — эктосильвиева ростральная борозда, 5 — супрасильвиева борозда, 6 — диагональная борозда, 7 — маргинальная (сагиттальная) борозда, 8 — венечная борозда, 9 — поперечная борозда, 10 — пресильвиева борозда, 11 – экто- и 12 — эндолатеральная борозды, 13 — крестовидная борозда, 14 — щель гиппокампа, 15 — борозда мозолистого тела, 16 – борозда валика, 17 — надваликовая борозда, 18 — борозда колена, 19 — шпорная борозда, 20 — надколенная борозда, 21 — свод, 22 — извилина гиппокампа, 22’ — грушевидная доля, 22” — поясная извилина, 23 — зрительный бугор, 24 — коллатеральная борозда.
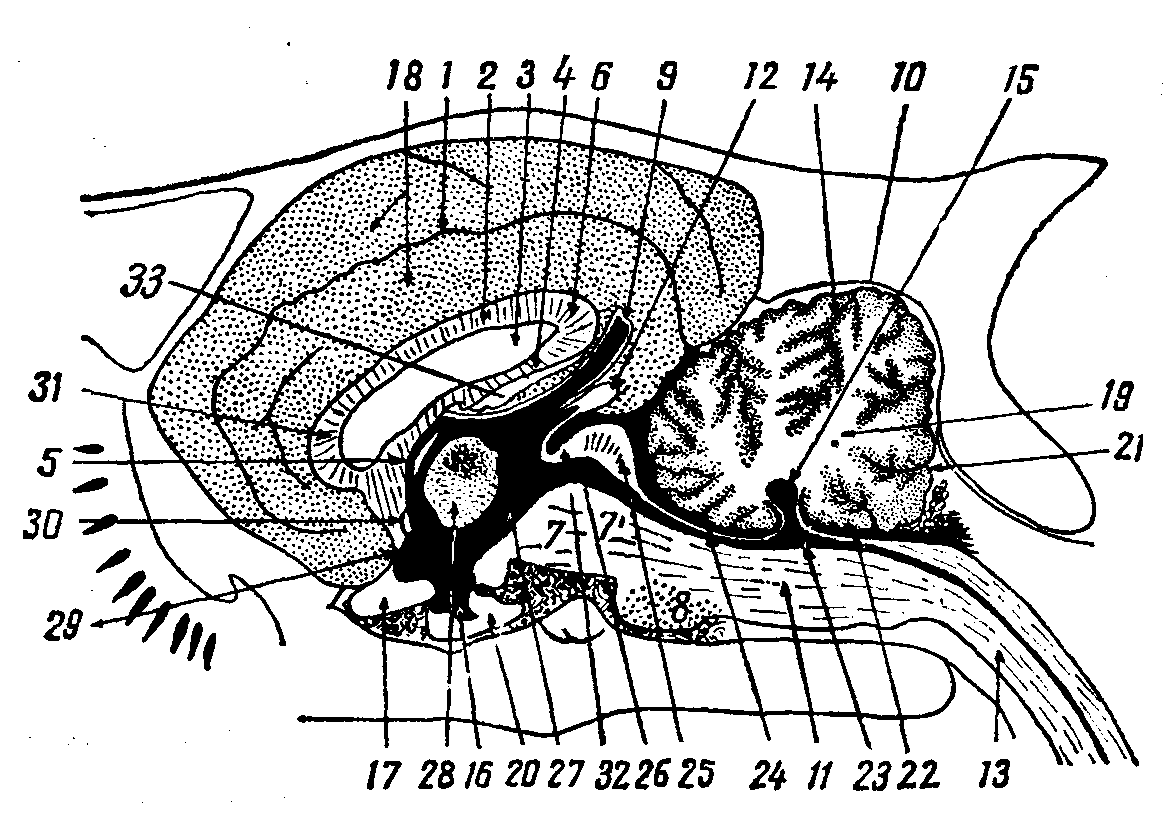
Рис. 14. Сагиттальный разрез головного
мозга:
1 — борозда валика, 2 — мозолистое тело, З — прозрачная межжелудочковая перегородка,4 — свод, 5 – межжелудочковое отверстие, 6 — валик мозолистого тела, 7 — ножка большого мозга, 7’ — чепец, 8 — мозговой мост, 9 — надшишковидный карман, 10 — мозжечок, 11 — продолговатый мозг, 12 — шишковидная железа, 13 – спинной мозг и центральный спинномозговой канал, 14 — первичная щель мозжечка, 15 — верхушка шатра, 16 — серый бугор и воронка, 17 — зрительный перекрест, 18 — поясная извилина, 19 — белое мозговое вещество червячка, 20 — гипофиз, 21 — предпирамидальная щель мозжечка, 22 — каудальный мозговой парус, 23 – четвертый мозговой желудочек, 24 — назальный мозговой парус, 25 — четверохолмие, 26 — мозговой водопровод, 27 — третий мозговой желудочек, 28 – зрительные бугры, 29 – концевая пластинка, 30 — ростральная комиссура, 31 — колено мозолистого тела,32 — каудальная спайка, 33 — спайка гиппокампа
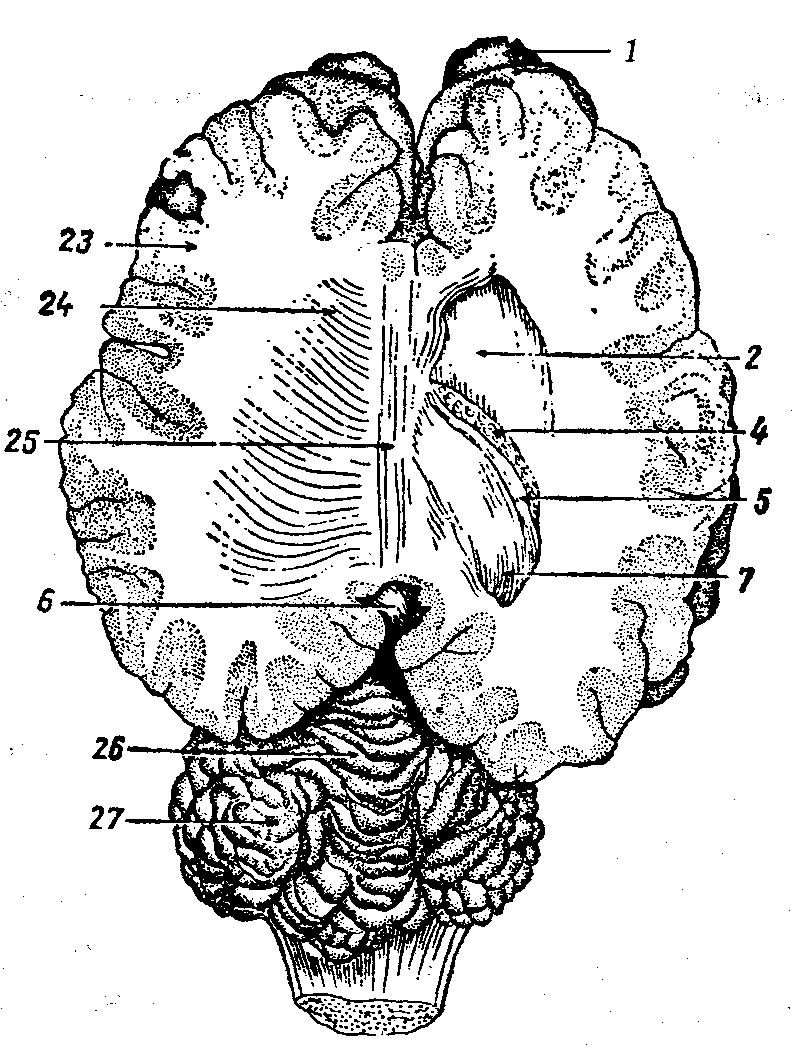 Рис.
15.
Горизонтальный срез полушарий большого
мозга лошади ( правый боковой желудочек
вскрыт):
Рис.
15.
Горизонтальный срез полушарий большого
мозга лошади ( правый боковой желудочек
вскрыт):
См. обозначения на рисунке 16.
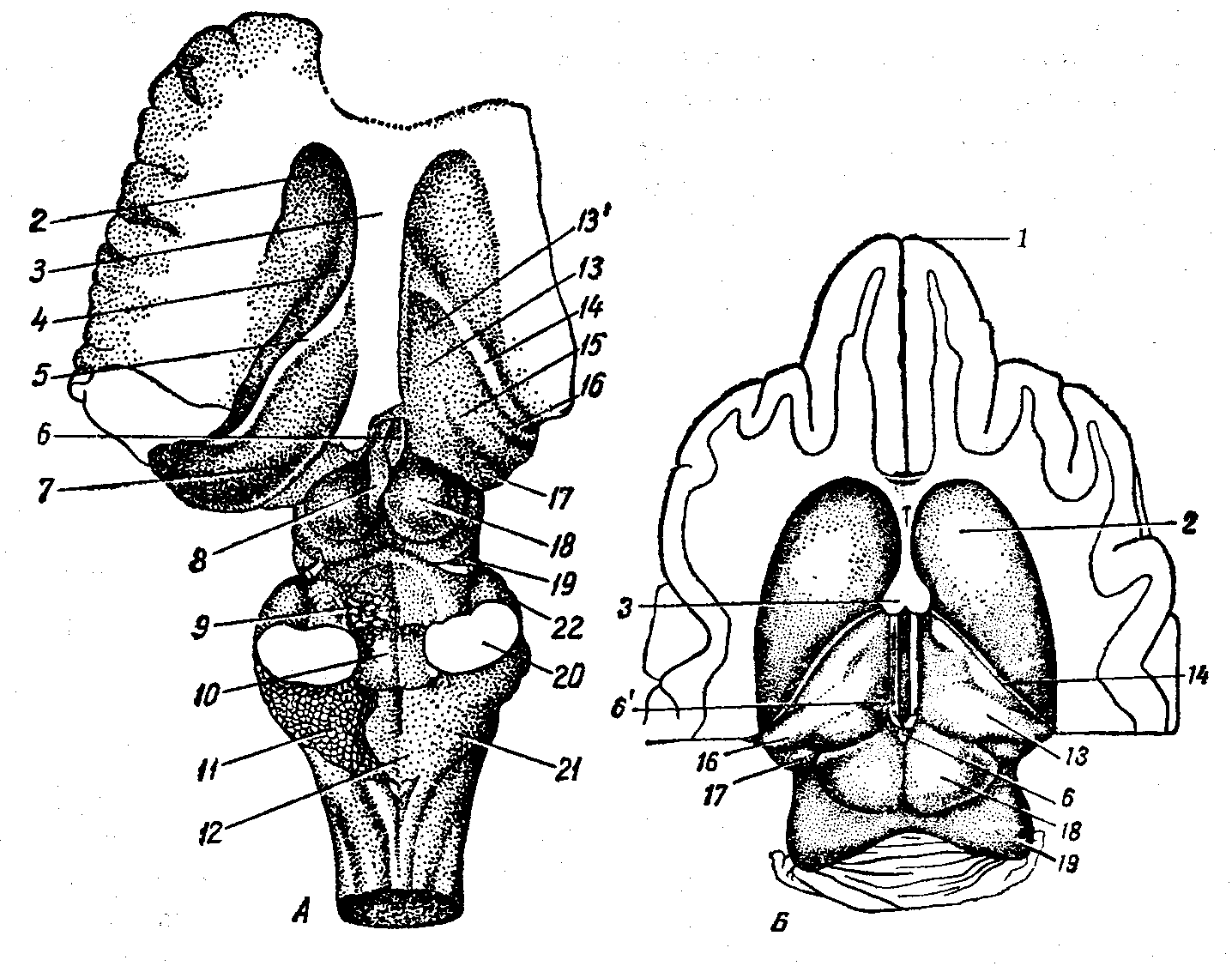
Рис. 16. Дно боковых желудочков большого мозга:
А — мозг лошади (слева задняя часть плаща срезана, чтобы показать гиппокамп — аммонов рог, справа гиппокамп удален, чтобы показать зрительный бугор). Б — мозг собаки после удаления задней половины плаща з гиппокампами; 1 — обонятельные луковицы, 2 — хвостатое ядро, З — свод, 4 – сосудистое сплетение бокового желудочка, 5 — бахрома, 6 — эпифиз, 6’ — уздечка, 7— желобоватый листок гиппокампа, 8 — надшишковидный карман, 9 — назальный мозговой парус, 10 — дно IV мозгового желудочка, 11 — сосудистое сплетение четвертого желудочка, 12 — каудальный мозговой парус, 13 — зрительный бугор, 13’ — его назальный бугорок, 14 — пограничная борозда, 15 — латеральный бугорок зрительного бугра, 16 — латеральное и 17 — медиальное коленчатые тела, 18 — ростральные и 19 — каудальные холмы четверохолмия, 20 — латеральные ножки мозжечка, 21 — каудальные ножки мозжечка, 22 — ростральные ножки мозжечка, 23 — белое мозговое вещество, 24 — лучистость спайки, в правой половине удалена, 25 — мозолистое тело, 26 — червячок, 27 — полушарие мозжечка
1) Обонятельная луковица — bulbus olfactorius — (рис. 12, 1) — парное образование в виде довольно плоского, вытянутого и загнутого дорсально мозгового отростка, который выдается за передний край полушария мозга в обонятельную ямку решетчатой кости. Дорсо-медиальный отдел луковицы построен из серого мозгового вещества, а латеро-вентральный — из белого мозгового вещества. В луковице находится желудочек обонятельной луковицы — ventriculus bulbi olfactorii; он является продолжением бокового желудочка мозга.
В обонятельную луковицу входит обонятельный нерв — n. olfactorius (1 пара). Он содержит многочисленные пучки нервных волокон — fila olfactoria, направляющиеся от обонятельных клеток слизистой оболочки носа к нервным клеткам луковицы. Таким образом, обонятельные луковицы являются первичными обонятельными центрами. (Нервный центр, по И. П. Павлову, «скопление и сцепление нервных клеток»).
2) От нервных клеток обонятельной луковицы начинаются обонятельные проводящие пути.
Они образуют белое мозговое вещество самой луковицы и обонятельные тракты — общий, медиальный и латеральный — tractus olfactorius communis, medialis et lateralis (2, 2, 2”). Латеральный обонятельный тракт переходит на грушевидную долю, покрывая на всем своем пути латеральную обонятельную извилину — gyrus olfactorius lateralis. Медиальный обонятельный тракт достигает медиальной поверхности плаща, формируя околообонятельное поле — area parolfactoria, а по пути покрывает медиальную обонятельную извилину — gyrus olfactorius medialis. Обонятельные тракты ограничивают обонятельньтй треугольник — trigonum olfactorium (3) — из серого мозгового вещества; они проводят импульсы от обонятельной луковицы к клеткам вторичных обонятельных центров в обонятельных извилинах, обонятельных треугольниках, околообонятельньих полях, в грушевидных долях, а также в гипоталамус и в средний мозг.
3) Грушевидяая доля — lobus piriformis (крючок — uncus) (б) — расположена медиально от латерального обонятельного тракта и каудально от обонятельного треугольника; медиально граничит с ножками большого мозга. Каудо-медиальной границей грушевидной доли является медиальная пограничная щель, или щель гиппокампа — fissura hippocampi (щель гиппокампа можно видеть на медиальной поверхности полушарий только после удаления промежуточного мозга) (рис. 16, 14;17, 9). В грушевидной доле заключена полость, представляющая собой задний отдел бокового желудочка мозга.
Грушевидная доля каудально переходит без четкой границы в извилину гиппокампа — gyrus hippocampi, расположенную на медиальной поверхности полушария, кзади и латерально от щели гиппокампа.
Извилина гиппокампа дорсально продолжается в поясную извилину — gyrus cinguli (рис.13, 22). Последняя, как часть плаща, проходит непосредственно дорсально от мозолистого тела и, огибая его спереди, соединяется с околообонятельным полем и представляет собой часть коры плаща (neopallium).
Грушевидная доля является вторичным обонятельным центром.
4) Гиппокамп, или аммонов рог — hiрроcampus, s. cornu ammonis — своим дорсальным отделом образует дно бокового желудочка мозга, позади хвостатого ядра, от которого отделяется сосудистым сплетением бокового желудочка. Гиппокамп представляет собой складку коры мозга в области щели гиппокампа и грушевидной доли. Он серповидно изгибается латеро-каудально и вентрально и теряется в стенке грушевидной доли. Аммоновы рога лежат дорсально на зрительных буграх, будучи отделены от них сосудистым сплетением третьего мозгового желудочка. Являясь высшими ассоциационными подкорковыми обонятельными и вкусовыми центрами, аммоновы рога связаны с различными участками коры больших полушарий и подкорковыми ядрами. Проводящие пути их образуют свод и его производные.
5) Свод — fornix, s. tractus hippocampo-mamillaris — содержит в своем составе проводящие пути, соединяющие гиппокамп с сосцевидным телом промежуточного мозга (рис. 14, 4 и 16, 3). Отдельные участки этого пучка волокон образуют желобоватый листок, кайму аммонова рога, ножки, столбы, тело и ростральную спайку мозга.
Желобоватый листок — alveus hippocampi — покрывает гиппокамп с его поверхности, обращенной в боковой желудочек мозга (рис. 15, 7). Он образован нервными волокнами, происходящими из серого мозгового вещества грушевидной доли и гиппокампа. По дорсо-латеральному краю эти волокна создают кайму гиппокампа — fimbria hippocampi, продолжающуюся назально в ножку свода — сrus fornicis. Последняя, соединяясь с ножкой другой стороны, становится коротким телом свода — соrpus fornicis. Тело свода служит дорсальной стенкой третьего мозгового желудочка; назально оно разделяется на два столба свода — columnae fonicis. Последние медиально от хвостатых ядер направляются к сосцевидному телу и серому бугру гипоталамуса (часть промежуточного мозга). Комиссура свода — соmissura fornicis — образована поперечными волокнами между ножками свода; она соединяет дорсальные концы аммоновых рогов друг с другом.
6) Между ростральным концом мозолистого тела (клювом) и столбами свода находится передняя спайка мозга — сomissura rostralis (рис. 14, 30). Она соединяет друг с другом обонятельный мозг обоих полушарий.
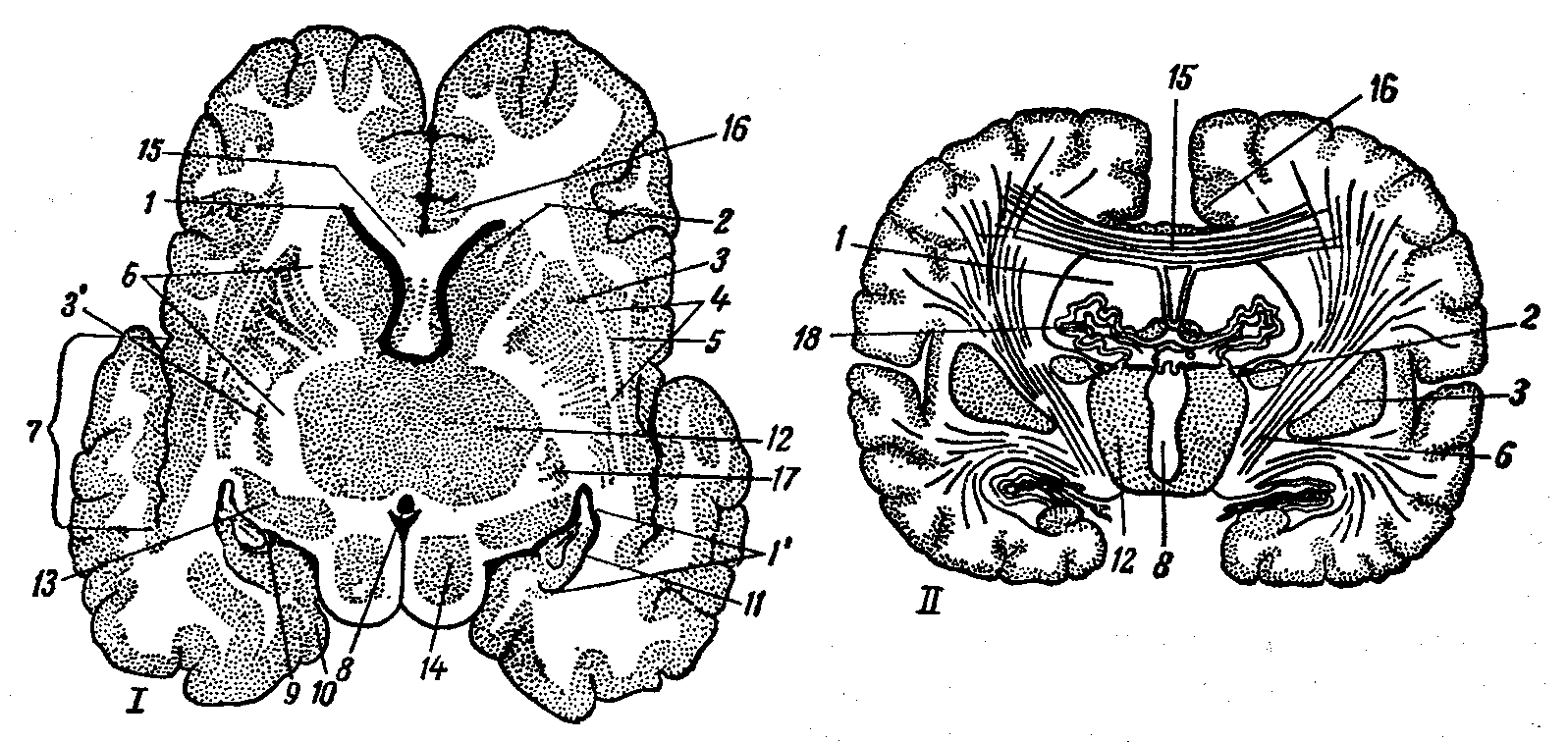
Рис. 17. Полосатое тело на разрезе:
I — горизонтальный срез большого мозга, II — поперечный срез мозга; 1 — полость бокового мозгового желудочка, 1’ — вентральный рог бокового желудочка, 2 — хвостатое ядро, 3 — чечевицеобразное ядро, 3' — бледное ядро, 4 — наружная капсула, 5 — ограда, 6 — внутренняя капсула, 7 — островок, 8 — третий мозговой желудочек, 9 — щель гиппокампа, 10 — грушевидная доля, 11 — гиппокамп, 12 — зрительные бугры, 13 — ганглий латерального коленчатого тела, 14 — четверохолмие, 15 — мозолистое тело и лучистость мозолистого тела,
16 — поясная извилина, 17 — миндалевидное тело, 18 — сосудистое сплетение боковых желудочков.
В) Полосатое тело — соrpus striatum — расположено впереди зрительных бугров, дорсально от обонятельных треугольников, состоит из четырех ядер: хвостатого, чечевицеобразного, миндалевидного тела и ограды. Между ядрами находятся внутренняя и наружная капсулы, образованные проекционными проводящими путями (рис. 16, 4-6).
На хвостатом ядре — nucleus caudatus — различают головку — сарut — и хвост — саuda. Головка хвостатого ядра своей дорсальной поверхностью образует дно бокового желудочка, назо-латерально от гиппокампа.
Чечевицеобразное ядро — nucleus lentiformis — располагается латерально от головки хвостатого ядра и зрительного бугра и отделяется от них внутренней капсулой — сарsula interna. Латеральная часть чечевицеобразного ядра называется скорлупой — putamen. Медиальная часть чечевицеобразного ядра более древнего происхождения, называется бледным ядром — globus pallidus. Оно связано с обонятельным мозгом, лежит латерально от зрительного бугра и впереди от ядра латерального коленчатого тела — ganglion deniculatum laterale. Латерально от чечевицеобразного ядра находится наружная капсула — сарsula externa, а латерально от капсулы — другое ядро в виде узкой полоски — ограда — claustrum. На границе между оградой, скорлупой и аммоновым рогом располагается миндалевидное тело — corpus amygdaloideum, которое является подкорковым обонятельным центром (рис. 17, 17).
Полосатые тела соединяются проводящими путями: 1) с корой большого мозга; 2) со зрительными буграми и гипоталамусом; 3) с ядрами сетчатого образования среднего мозга — nucleus motorius tegmenti (красное ядро и др.), с ядрами моста и продолговатого мозга (каудальные оливы); 4) с ядрами черепномозговых нервов.
Через полосатые тела замыкаются различные рефлекторные цепи: а) периферический рецепторный аппарат — зрительные бугры — полосатые тела — соматические и висцеральные эффекторные аппараты; б) кора — полосатое тело — соматические и висцеральные эффекторные аппараты. У млекопитающих ядра полосатых тел являются важнейшими подкорковыми двигательными центрами: 1) координированных непроизвольных движений (ходьба, бег, лазание); 2) регуляции мышечного тонуса в состоянии покоя и движения; 3) безусловных рефлексов (в виде жестов у человека, позы и мимики) и, наконец, высшими подкорковыми вегетативными центрами. Полосатые тела функционируют как единое целое, но отдельные их части действуют противоположно: например, полосатое тело тормозит движения, а. globus pallidus (бледное ядро) вместе с медиальным ядром зрительного бугра, напротив, усиливает их.
Фило- и онтогенез. Из более древних передней и вентральной стенок конечного мозга — telencephalon — происходят обонятельные доли и полосатые тела, а из более поздней дорсальной его стенки — плащ (рис. 18).
I. Развитие обонятельных долей мозга обусловливается наличием обонятельного анализатора, который у водных животных имеет исключительное значение при ориентации во внешней среде. Отростки чувствительных обонятельных клеток заканчиваются в обонятельных луковицах — bulbus olfactorius, составляющих часть обонятельных долей — lobi olfactorii — конечного мозга, проксимальные отделы которых формируют древнейшую обонятельную кору — paleocortex — из серого мозгового вещества. У млекопитающих она образует обонятельные извилины и обонятельные треугольники — trigonum olfactorium, являющиеся вторичными обонятельными центрами (1).
II В вентральной стенке концевого мозга, дорсально от обонятельной коры — paleocortex — очень рано появляется крупноклеточный базальный ганглий — ganglion basale, s. paleostriatum (1”). Это высший двигательный центр. Базальный ганглий сохраняется и у млекопитающих животных в виде бледного ядра — globus pallidus (рис. 17, 3’). Позднее у наземных животных разрастаются дополнительные мелкоклеточные ядра, образующие в целом neostriatum — скорлупы (начиная с рептилий), а у млекопитающих еще хвостатого ядра — nucleus caudatus — (2).
Кроме того, обособляются новые ядра — ограда и миндалевидное ядро — claustrum et nucleus amygdalae. С появлением вторичного плаща (у млекопитающих) сложный базальный ганглий пронизывается внутренней и наружной капсулами — сарsula interna et externa — из проводящих путей, идущих в кору плаща и обратно (4, 6). В силу зтого и возник новый термин — полосатое тело — соrpus striatum.
III. В эволюции плаща наблюдаются две формации различной функции и строения, не считая мембранообразного примитивного плаща — раllium membranaceum, характерного для водных животных и состоящего из эпендимы (рис. 18, 6). У наземных животных сначала возникает кора более простой, трехслойной, клеточной структуры. Из нее формируются высшие обонятельные центры: грушевидные доли, извилины гиппокампа и их производные. Все эти части объединяются понятием первичный плащ — аrhipallium (аrchicortex — старая кора) (рис. 344, 6’) (или грушевидвая область плаща – аrea piriformis).
Первичный плащ характеризуется расположением серого мозгового вещества под белым мозговым веществом. Архипаллиум, так же как и палэопаллиум, лучше всего выражен у макросматических животных — macrosmates, обладающих острым обонянием, в отличие от микросматов — microsmates, у которых орган обоняния менее развит (приматы и некоторые другие).
У млекопитающих на границе между обонятельной долей — lobus olfactorius — и первичным плащом — archipallium — появляется вторичный плащ — neopallium (neocortex — новая кора) (рис. 18, 6”). Вторичный плащ характеризуется поверхностным расположением серого мозгового вещества в виде коры над белым мозговым веществом. Зачатки его появляются еще у рептилий. По мере своего разрастания новый плащ оттесняет архипаллиум в медиальную сторону и вентрально; при этом в области щели гиппокампа образуется складка коры в виде гиппокампа, а сам неопаллиум занимает всю дорсо-латеральную часть полушария и, распространяясь каудально, покрывает промежуточный и даже средний мозг с их дорсальной и боковых поверхностей (рис. 22).
Разрастание вторичного плаща вызывается внедрением в него новых проекционных нервных волокон из промежуточного мозга, являющихся проводниками различных анализаторов — кожного, зрительного, слухового, мышечного, и развитием новых ассоциационных нейронов и центров, включая formatio reticularis.
Кора вторичного плаща в ряду животных чрезвычайно усложняется в своем строении, резко дифференцируются ее функции и структура и увеличиваются размеры. У крупных млекопитающих животных плащ обычно несет извилины и борозды. Такие животные называются gyrencephali. Извилины в ряду животных располагаются неодинаково (рис. 13). В одних случаях (у хищных и копытных) они в основном идут дугами вокруг поперечной сильвиевой борозды, у приматов извилины образуют две системы — лобвую и теменную. Обе системы разделены сильвиевой бороздой. У третьей группы животных сильвиева борозда отсутствует, борозды идут в передней части мозга продольно, а в задней поперечно. В силу сказанного абсолютная гомологизация извилин между животными разных отрядов крайне затруднительна, а в отдельных случаях может быть и невозможна (макроскопически).
У мелких животных извилины вообще отсутствуют (животные с гладким головным мозгом — lissencephali). Наибольшее число извилин насчитывается у слонов и китов (а в ряду приматов — у человека). В онтогенезе извилины появляются также не сразу, а в определенной последовательности.