

I етап дихання. Гліколіз.
Гліколіз - процес анаеробного розпаду глюкози, що проходить із вивільненням Е, кінцевим продуктом якого є піровиноградна к-та (ПВК). Гліколіз - загальний спільний етап аеробного дихання і всіх видів бродіння.
Реакції гліколізу протікають у розчинній частині цитоплазми. В цитозолі гліколітичні ферменти організовані в мультиензимні комплекси на активних філаментах цитоскелету.
Біохімія процесу. Ланцюг реакцій гліколізу можна розбити на три етапи.
1. Підготовчий етап - фосфорилювання гексози і її розщеплення на дві фосфотріози. Глюкоза (піранозна форма молекули) фосфорилюється АТФ з участю гексокінази, перетворюючись у глюкозо-6-фосфат, який ізомеризується глюкозофосфатізомеразою у фруктозо-6-фосфат. Цей перехід необхідний для утворення більш лабільної фуранозної форми молекули гексози. Фруктозо-6-фосфат фосфорилюється за участю АТФ фосфофруктокіназою. В результаті утворюється фруктозо-1,6-дифосфат.
Фруктозо-1,6-дифосфат – лабільна форма із симетрично розміщеними фосфатними групами, що несуть від'ємний заряд і відштовхуються один від одного електростатично. Така структура легко розщеплюється альдолазою на 3-фосфогліцериновий альдегід (3ФГА) і фосфодіоксіацетон (ФДА), які легко перетворюються один в одного (тріозофосфатізомераза).
2. Перше субстратне фосфорилювання, яке починається з 3ФГА і закінчується 3-фосфогліцериновою кислотою (3ФГК).
Фермент дегідрогеназа фосфогліцеринового альдегіду (НАД-залежний SН-фермент) утворює з 3ФГА фермент-субстратний комплекс, в якому відбувається окислення субстрату та передача е- і Н+ на НАД+. При цьому утворюється АТФ.
Таким чином, в результаті 2-го етапу гліколізу утворюються на кожну тріозу 1 мол АТФ і відновлений НАДН.
3. Завершальний етап. 3ФГК з допомогою фосфогліцератмутази перетворюється в 2ФГК. А далі енолаза каталізує відщеплення молекули води від 2ФГК і утворюється фосфоенолпіруват – сполука, яка містить високоенергетичний фосфатний зв'язок. Цей фосфат при участі піруваткінази передається на АДФ, а енолпіруват переходить у більш стабільну форму – піруват.
Енергетичний вихід гліколізу. На 2-му і 3-му етапах утворюється 4 мол. АТФ і 2 мол. НАДН, на першому етапі витрачається 2 мол. АТФ. Окислення 1 мол. НАДН = синтезу 3-х мол. АТФ. Таким чином, у процесі гліколізу утворюється 8 мол. АТФ.
Вільна Е гідролізу 1 мол АТФ = 41, 87 кДж/моль (10 ккал), 8 мол. АТФ дають 335 кДж/моль (80 ккал)
Функції гліколізу в клітині:
- здійснює зв'язок між субстратами дихання й циклом Кребса;
- постачає на потреби клітини АТФ і НАДН;
- продукує інтермедіати, необхідні для синтетичних процесів у клітині.
II етап дихання. Цикл Кребса
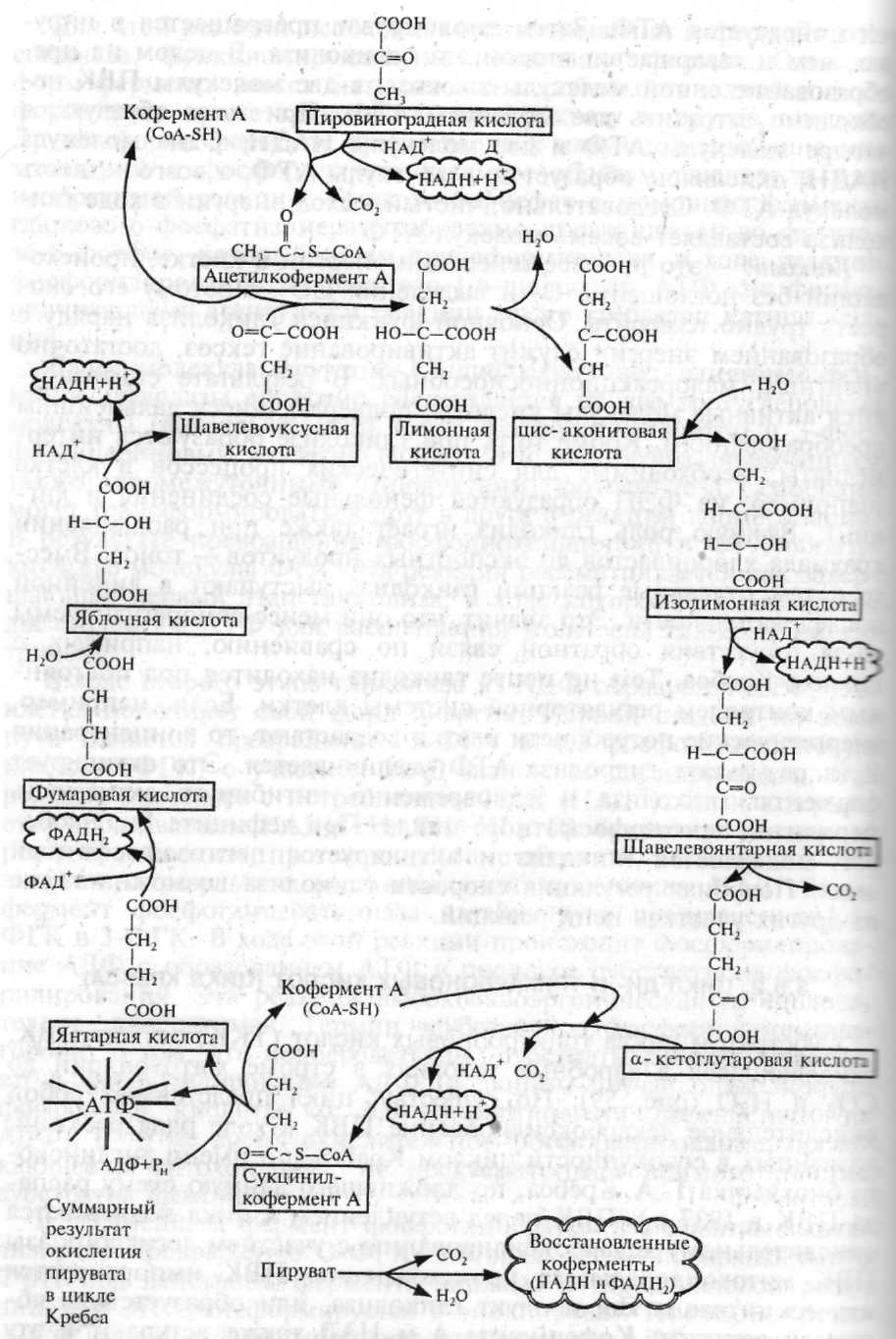
У 1937 р. англійським біохіміком Г.А.Кребсом була запропонована схема окислення ди- і трикарбонових кислот до СО2 через ”цикл лимонної кислоти” за рахунок віднімання водню. Таким чином на тваринних об'єктах був відкритий цикл Кребса, а в 1939 р. англійським дослідником А.Чібнелом була доказана його наявність у рослин.
В циклі Кребса при наявності кисню піруват повністю окислюється до СО2 і Н2О. Усі ділянки цього процесу локалізовані в матриксі або на внутрішній поверхні мітохондрій. Виділяють 8 етапів.
1) Безпосередньо в циклі окислюється не сам піруват, а його похідне – ацетил-СоА. Тому перший етап – це утворення активного ацетилу в ході окисного декарбоксилювання. Цей процес здійснюється при участі піруватдегідрогеназного мультиферментного комплексу (входять 3 ферменти і 5 коферментів). В ході складних перетворень через проміжні сполуки з коферментами з ПВК утворюються ацетил-СоА (із високоенергетичним тіоефірним зв'язком), СО2 і НАДН.
2) Цикл Кребса починається із взаємодії ацетил-СоА з енольною формою щавлево-оцтової кислоти, які при дії цитратсинтази перетворюються в лимонну к-ту. При цьому витрачається енергія тіоефірного зв'язку.
3) Наступний етап циклу включає дві реакції і каталізується ферментом аконітатгідратазою - в результаті синтезується ізолимонна кислота.
4) Ізолимонна к-та під дією НАД-залежної ізоцитратдегідрогенази окислюється в нестійку сполуку – щавлевобурштинову к-ту, яка декарбоксилюється з утворенням α-кетоглутарової кислоти.
5) α-кетоглутарат піддається реакції окислювального декарбоксилювання → α-кетоглутарат-дегідрогеназний мультиферментний комплекс — в результаті чого виділяється СО2, утворюється НАДН і сукциніл-СоА – високоенергетичний тіоефір.
5/) При участі сукциніл-СоА-синтетази із сукцинілу-СоА, АДФ і Н3РО4 утворюється бурштинова к-та, АТФ (субстратне фосфорилювання), регенерує молекула СоА.
6) Далі бурштинова к-та (сукцинат) окислюється до фумарової.
7) Фумарова к-та приєднуючи Н2О перетворюється в яблучну (малат).
8) Яблучна з допомогою малатдегідрогенази окислюється в щавлевооцтову, яка самочинно переходить в енольну форму, реагуючи з черговою молекулою ацетил-СоА і цикл повторюється.
На протязі одного обороту циклу при окисленні пірувату виділяється 3 молекули СО2, відбувається включення 3-х молекул Н2О і видалення 5-ти пар атомів водню.
Значення та енергетичний вихід
Цикл Кребса відіграє дуже важливу роль в обміні речовин рослинного організму. Він служить кінцевим етапом окислення не лише вуглеводів, але й білків, жирів та ін. сполук. В ході реакцій циклу звільняється основна кількість енергії, яка міститься в субстраті, і більша її частина утилізується у високоенергетичних фосфатний зв’язках.
В ході окислення пірувату утворюються 3НАДН [1, 5, 8], НАДФН [4] і ФАДН2 [6] (=2АТФ), +1 мол. АТФ при субстратному фосфорилюванні (5/) = всього 15 молекул АТФ, а для 2-х молекул ПВК=30 молекул АТФ.
Отже, при окисленні глюкози в процесі дихання при функціонуванні гліколізу й циклу Кребса утворюється 38 молекул АТФ, що складає 380 ккал/моль.
Значення Циклу Кребса цим не вичерпується. Багато проміжних продуктів циклу використовується для синтезу різноманітних сполук (амінокислот, жирів, вуглеводів, поліізопренів і т. д. )