

39. Аеробне окислення глюкози. Етапи перетворення глюкози до со2 і н2о.
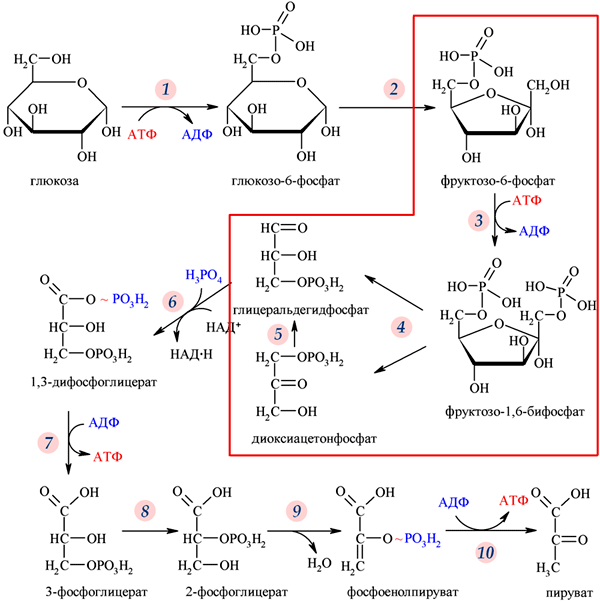
Для більшості тканин людини та вищих тварин в умовах нормальної життєдіяльності характерний аеробний гліколіз, тобто утворення з глюкози пірувату, який у подальшому окислюється до вуглекислого газу й води. Анаеробний гліколіз має місце переважно в м`язах при інтенсивній фізичній діяльності, тобто при відносній кисневій недостатності, та в деяких високоспеціалізованих клітинах (зокрема, в еритроцитах, в яких відсутні мітохондрії) або за певних патологічних умов (клітини злоякісних пухлин). Реакції гліколізу перебігаютьу цитозолі клітини і каталізуються ферментами, що локалізовані в цьому компартменті. Виділяють дві стадії гліколізу: 1. Розщеплення молекули глюкози до двох молекул фосфотріоз (гліцеральдегід-3-фосфату та діоксіацетонфосфату). Ця стадія включає в себе послідовність реакцій, які потребують витрати двох молекул АТФ на кожну молекулу глюкози, що розщеплюється. 2. Перетворення двох молекул фосфотріоз на дві молекули пірувату (або лактату). Ця стадія включає в себе окислювально-відновлювальні реакції ("гліколі-тична оксидоредукція"), які супроводжуються генерацією чотирьох молекул АТР. Таким чином, у результаті розщеплення однієї молекули глюкози в реакціях аеробного або анаеробного гліколізу сумарний вихід АТР складає дві молекули, що можна подати таким рівнянням: D-глюкоза + 2 АДФ + 2 Фн ? 2 піруват (лактат) + 2 АТФ
40. Окислювальне декарбоксилювання пірувату. Ферменти, коферменти та послідовність реакцій в мультиферментному комплексі.
Окислювальне декарбоксилювання пірувату каталізується піруватдегідрогеназним комплексом — мультиферментною системою, яка в клітинах еукаріотів міститься в мембранах мітохондрій, а у прокаріотів — у цитоплазмі. До складу цього комплексу входять три ферменти, що каталізують три послідовні стадії перетворення пірувату на ацетил-КоА: • піруватдегідрогеназу [КФ 1.2.4.1]; • дигідроліпоїлацетилтрансферазу [КФ 2.3.1.12]; • дигідроліпоїлдегідрогеназу [КФ 1.8.1.4] і п'ять коферментів і простетичних груп: • тіаміндифосфат (ТДФ, або ТПФ); • ліпоєву кислоту(ЛК); • КоА; • ФАД; • НАД+ У результаті цього процесу утворюється ацетилкоензим А — основний субстрат окислення в циклі трикарбонових кислот — та відновлена форма НАД+. Сумарне рівняння окислювального декарбоксилювання пірувату: CH-CO-COOH + НАД+ + КоА-SH -> CH-CO-КоА + НАДН + Н+ + С02 Ферментативні стадії утворення ацетил-КоА з пірувату: Стадія I (декарбоксилювання)— каталізується піруватдегідрогеназою (Е1), коферментом якої є ТДФ. На цій стадії відбувається взаємодія пірувату з С-2 тіазольного кільця молекули тіаміну; в результаті реакції утворюється зв’язаний із ферментом гідроксиетильний похідний тіаміндифосфату: Стадія II— каталізується центральним ферментом комплексу дигідроліпоїлацетилтрансферазою (E2), яка переносить гідроксиетильну групу від ТДФ(Е1) на простетичну групу ферменту E2, що є окисленою формою ліпоєвої кислоти (ЛК); в результаті реакції утворюється ацетилтіоефір відновлених ліпоїльних груп ферменту E2, що містить макроергічний зв’язок: Стадія ІІІ— дигідроліпоїлацетилтрансфераза переносить ацетильну групу від відновленої ліпоєвої кислоти на коензим А: Стадія IV — окислення відновленої форми ферменту Е2 ФАД-залежною дигідроліпоїлдегідрогеназою (E3): Стадія V — перенесення атомів водню від відновленої ФАД-групи дигідроліпоїлдегідрогенази на НАД+ з утворенням НАДН: Відновлений НАДН, що утворюється в результаті окислювального декарбоксилювання пірувату, в аеробних умовах окислюється в мітохондріальному електронотранспортному ланцюзі з генерацією шести (2x3) молекул АТФ. Біологічна роль окиснювального декарбоксилювання пірувату полягає: 1) в катаболізмі пірувату до одного з кінцевих продуктів - С02 (виводиться з організму або використовується для синтезу), 2) в утворенні макроергічної сполуки - ацетил-Ко А (піддається подальшому окисненню в циклі трикарбонових кислот або використовується в реакціях анаболізму); 3) в синтезі відновленого еквіваленту - НАДН, який окиснюється в дихальному ланцюгу мітохондрій. Реакції окиснювального декарбоксилювання пірувату регулюються в організмі на рівні піруватдегідрогенази: 1) алостерично (надлишок продуктів реакції ацетил-КоА НАДН інгібує фермент, а проміжний продукт гліколізу фруктозо-1,6-дифосфат, НАД+, КоА - є активаторами піруватдегідрогенази); 2) шляхом хімічної модифікації - фосфорилування. Фосфорильована форма піруватдегідрогенази є неактивною, а дефосфорильована — активна. Тому інсулін підвищує активність комплексу, а глюкагон, адреналін (діють за аденілатциклазним каскадом) гальмують активність ферменту. Окислювальне декарбоксилювання пірувату – єдиний шлях його катаболізму, тому недостатність вітамінів (у перше чергу вітаміну В1) призводить до порушення процесу, зниження утворення АТФ і проявляється порушенням діяльності ЦНС. Піруватдегідрогеназна недостатність в організмі призводить до підвищення концентрації лактату, пірувату, аланіну, що супроводжується ацидозом.