

Кайданов Генетика популяций
.pdfУже через 10 поколений отбора частота рецессивных гомозигот падает примерно в 20 раз по сравнению с исходным уровнем. Частота выщепления белых телят составляет только 0,5%. Теперь для выполнения поставленной задачи можно рекомендовать селек ционерам начать частичную выбраковку и гетерозиготных, чалых телят.
Чем меньше становится частота рецессивной аллели, тем менее эффективным оказывается отбор, если элиминации подвергаются только рецессивные гомозиготы. В рассматриваемом примере q100= 0, отбора частота рецессивных гомозигот падает в 300 раз от первона чальной, то за последующие 50 поколений только в 3 раза. Графи чески подобная зависимость отражена на рис. 4.2.
При низких частотах рецессивной аллели она будет представлена в популяции преимущественно в гетерозиготном состоянии. Следо вательно, элиминации будут подвергаться лишь редкие гомозигот ные носители этой аллели и генотипическая структура популяции,
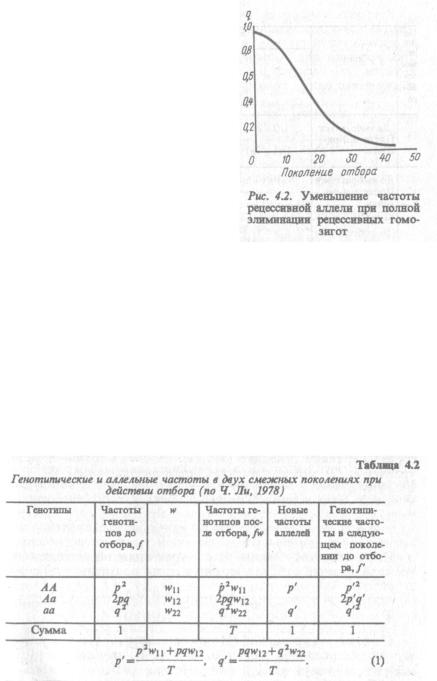
как видно из рис. 4.2, будет меняться очень слабо.
Эффективность действия отбора при частичной элиминации рецессивных го
мозигот. Данная ситуация встречается значительно чаще, чем рассмотренная выше. Частичной элиминации в случае полного доминирования подвергаются гомозиготные носители рецессивных мутаций, снижающих жизнеспособ ность, плодовитость и другие приспо собительно важные свойства.
По-прежнему будут анализировать ся элементарные генетические измене ния в панмиктической популяции. Под элементарными генетическими измене ниями будем подразумевать сдвиги
в частотах пары аллелей одного гена в смежных поколениях.
Введем понятие относительной приспособленности, измеряемой величиной w. Эта величина характеризует относительный вклад в оставляемое потомство особей каждого из трех генотипических классов и может меняться в пределах от 0 до 1. В рассматриваемой ситуации доминантные гомозиготы и гетерозиготы обладают мак симальной адаптивной ценностью (w11 = w12=l), тогда как рецессив ные гомозиготы имеют показатель w, отличный от 0 и 1 (0 < W2 2 < 1). Если, скажем, W2 2 = 0,99, то это означает, что на каждые 100 потом ков, оставляемых доминантными особями, будет приходиться в сре днем 99 потомков, оставляемых рецессивными особями. Показа тель ^ = I - W будем называть коэффициентом отбора.
Занесем в табл. 4.2 частоты генотипических классов и частоты аллелей, измененные под действием отбора за одно поколение.





дов Н. В. Тимофеев-Ресовский, а также французские исследователи Ф. Л' Эритье и Ж. Тессье приступили к опытам с искусственными популяциями дрозофил, содержащимися в специальных популяци онных ящиках (рис. 4.3). В таких ящиках можно контролировать условия содержания мух (количество корма, температурный и све товой режим, влажность и др.), а также задавать определенные соотношения нормальных и мутантных особей.
На рис. 4.4 представлены результаты одного из экспериментов, в котором в исходном поколении к мутантным мухам ebony (е) с черной окраской тела были добавлены единичные особи дикого
типа. Видно, как в ряду поколений шло падение частоты мутантных особей, однако полностью они не исчезли. Спустя 20 поколений с на чала эксперимента частота рецес сивных гомозигот стабилизирова лась на уровне 8—9%, дальнейше го их уменьшения не наблюдалось. Специальная проверка показала, что гомозиготы е/е существенно уступают по жизнеспособности но рмальным гомозиготам + / + . Зато гетерозиготы +/е превосходят не только мутантных, но и нормаль ных гомозигот. Отрицательный от бор гомозигот компенсируется со временем положительным отбо ром гетерознгот. В основе стабили-

зации лежит механизм моногенного гетерозиса, или сверхдоминирования.
Детально изученным примером сверхдоминирования у человека на популяционном уровне может служить поддержание разных форм гемоглобина. Серповидно-клеточная анемия определяется аномальным строением гемоглобина (форма S), а именно заменой в В-цепи в 6-м положении валина на глутаминовую кислоту. Гете розиготные носители аллели HbS в обычных условиях клинически здоровы, но не могут находиться в условиях пониженного парци ального давления кислорода (например, в горах). У гомозигот по этой аллели с раннего детства развиваются расстройства кровооб ращения. В их основе лежит полимеризация молекул гемоглобина, сильное изменение формы эритроцитов (рис. 4.5), их гемолиз и рас пад.
Данная болезнь распространена в субтропиках и тропиках Аф рики и Евразиатского континента, при этом гетерозиготы по аллели
HbS невосприимчивы к другой тяжелой болезни — малярии. Там, где есть малярия, частота аллели HbS оказывается, как правило, высокой (рис. 4.5).
А. Аллисон в 1956 г. нашел в одном из округов Танзании следующие частоты генотипических классов в выборках из 654 взрослых и 287 детей (табл. 4.4). Данные таблицы указывают на повышенную смертность в ходе постнатального развития рецессивных гомозигот (SS) и на относительно высокую выживаемость! гетерозигот (AS). Эти данные позволяют оценить относительную приспособленность особей трех генотипических классов. Для оценки нужно соотнести генотипические частоты взрослых к таковым у де-1
AA |
AS |
SS |
тей. Тогда получим 0,929 |
1,228 0 |
242.Затем находим относитель- |
ные значения w, приняв W12 = 1 (табл. 4.4). Теоретически ожидаемые частоты аллелей в популяции в состоянии равновесия рассчитываются с помощью формул (7) или (8). Видно, что эти частоты весьма 1 близки к наблюдаемым.
Таблица 4.4
Частоты встречаемости в популяции человека (Танзания) особей трех генотипических классов (взрослых и детей), различающихся
по формам гемоглобина А и S (по A. Allison, 1956)
Частоты генотипов у |
AA |
AS |
SS |
q |
Взрослых |
0,612 |
0,381 |
0,007 |
0,198 |
Детей |
0,659 |
0,310 |
0,031 |
0,186 |
w |
0,757 |
1 |
0,197 |
|
|
|
|
|
q'= 0,233 |
s |
0,243 |
0 |
0,803 |
|
Таким образом, надежное сохранение популяции в условиях заражения малярийным плазмодием обеспечивается невосприим чивостью к нему гетерозиготных особей. Однако за это популяция! вынуждена расплачиваться низкой жизнеспособностью рецессивных гомозигот. Такая ситуация, которая сложилась за длительное время 1 естественно-историческим путем, была бы биологически оправданной для любого вида животных и растений и даже для человека в историческом прошлом. Но в настоящее время ее невозможно принять как неизбежное зло, поскольку жизнь каждого человека бесценна. К сожалению, эффективных методов лечения серповидноклеточной анемии пока еще не найдено.
Проблема гетерозиса на популяционном уровне с конца 30-х годов выдвинулась в генетике популяций на одно из ведущих мест. Об этом уже было упомянуто в вводной главе и подробно будет рассказано в гл. 5.
Устойчивость динамического равновесия аллельных частот. С по мощью рекуррентных (1) и разностных уравнений [по аналогии с (2),
