

Кайданов Генетика популяций
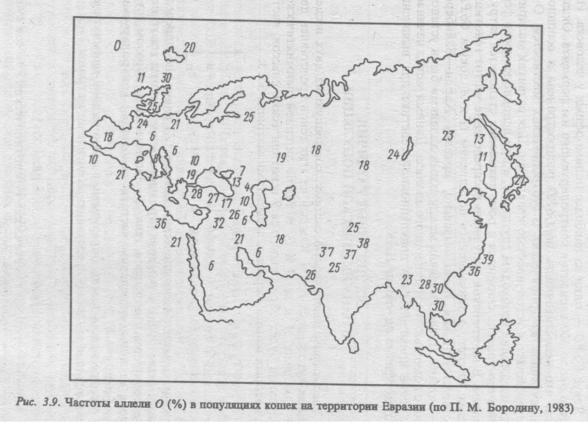
.pdfарабы и викинги, тогда как римляне имели, очевидно, иные при страстия. Во всяком случае там, где Римская империя некогда оказывала свое влияние, аллель О встречается относительно редко.
В 1930 г. Р. И. Серебровская выполнила популяционно-генети- ческое исследование дальтонизма, или цветовой слепоты (признак, сцепленный с полом), у жителей Москвы. В выборке из 5223 лиц мужского пола было выявлено 348 дальтоников (6,7%). Исходя из равновесного распределения в популяции частот генотипических классов можно утверждать, что частота рецессивных гомозигот соответствует частоте рецессивной аллели, т. е. q=0,067, а p = 0,933. Подставив эти значения в формулу Харди—Вайнберга, найдем вероятные частоты трех генотипических классов среди особей женс кого пола: p2 = 87,05%, 2pq= 12,50%, q2 = 0,45%.
Обратим внимание на соотношение частот рецессивных гемизигот и гомозигот. Это соотношение равно q/q 2 = l/q. Чем меньше q, тем в большей пропорции в панмиктической популяции будут пред ставлены рецессивные мужские особи по сравнению с женскими.
Врассмотренном примере это соотношение 1:15.
Учеловека в Х-хромосоме локализованы гены, мутации в кото рых влекут за собой такие аномалии, как гемофилия, дальтонизм, некоторые миопатии, ихтиоз (грубое строение кожи), болезни об мена. В сегменте Xq28, расположенном недалеко от дистального конца длинного плеча X-хромосомы, сосредоточен целый кластер

(набор) генов. Помимо уже упомянутых генов гемофилии и цвето вой слепоты здесь локализованы гены умственной отсталости, глю- козо-6-фосфатдегидрогеназы (G6PD) и ряд других.
Локус G6PD — один из наиболее полиморфных у человека. Из вестно около 300 вариантов контролируемого им фермента. Оказа лось, что недостаточность по G6PD распространена в основном в популяциях из тропических и субтропических местностей. Она сопровождается устойчивостью к малярии. Для развития малярий ного плазмодия необходим глутатион, а при неполноценности фер мента его уровень снижается и рост плазмодия становится затруд ненным. В нигерийской популяции распределение частот генотипических классов, различающихся по аллелям GnA локуса G6PD, соответствует, как было выяснено, правилу Харди—Вайнберга (L. Luzzatto, 1988). Различия между этими аллелями были установ лены с помощью рестрикционного анализа. Данный метод начинает все шире применяться в популяционной генетике для выявления полиморфизма на уровне ДНК.
3.7.Д И Г Е Н Н Ы Е Р А З Л И Ч И Я
ВП А Н М И К Т И Ч Е С К О Й П О П У Л Я Ц И И
Рассмотрим генетическую систему, состоящую из двух несцепленных аутосомных генов. Каждый из этих генов представлен дву мя аллелями: А и а; В и b. В такой дигенной панмиктической популяции будет образовываться 9 генотипических классов, часто ты которых можно выразить элементами матрицы 3x3:
AABB |
AABb |
ААbb |
Z1 1 |
Z1 2 |
Z1 3 |
AaBB |
AaBb |
Ааbb |
Z21 |
Z22 |
Z2 3 |
ааВВ ааВb аabb Z31 Z32 z33
Cумма частот генотипических классов (Z) равна, по определе нию, 1 (или 100%). Суммируя строчки, найдем частоты генотипов, контролируемых аллелями гена А. Аналогичным образом, сум мируя столбцы, определим частоты генотипов, контролируемых
аллелями гена В.
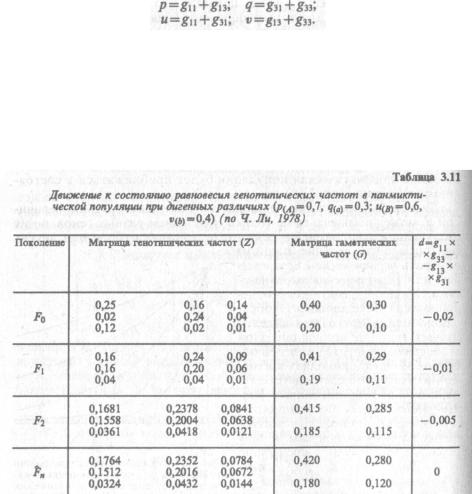
Четыре класса гамет, образуемых в дигибридной панмиктичес кой популяции, удобно представить элементами матрицы 2x2:
AB Ab G g2\11 g13 аВ ab _g31 g33
Между частотами гаметических и генотипических классов суще ствуют следующие зависимости:
Эти зависимости основаны на том, что гомозиготы дают все
гаметы одного класса, моногетерозиготы — гаметы двух |
классов |
в равной пропорции и, наконец, класс дигетерозигот |
образует |
С одинаковой частотой все четыре класса гамет. |
|
Обозначим частоты аллелей А и а как р и q, а частоты аллелей В и Ь — как u и v. После нахождения частот гаметических классов легко рассчитать частоты аллелей:
Генотипическая структура дочернего поколения может быть J установлена, если известны гаметические частоты в родительском поколении. Для ее установления достаточно подставить частоты гаметических классов в решетку Пеннета, произвести перемножение и затем просуммировать частоты одинаковых генотипических классов.
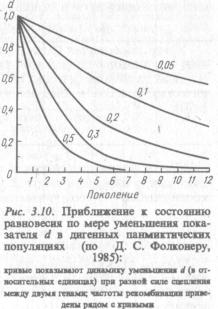
В табл. 3.11 представлены частоты генотипических и гаметических классов в последовательном ряду поколений панмиктической популяции при дигенных различиях.
Можно убедиться, проведя суммирование по строчкам и сто лбцам, что уже после первого поколения случайных скрещиваний распределение генотипических частот по каждой паре аллелей в от дельности становится равновесным:
|
AA |
Aa |
|
aa |
|
BB |
Bb |
|
bb |
p2 |
= 0,49 |
2pq=0,42 |
q2 |
= О,О9 |
u2 |
= 0,36 2uv; = 0,48 |
v2 |
=0,16. |
|
Однако в целом с учетом различий по обеим парам аллелей матрица генотипических и гаметических частот в ряду поколений будет продолжать изменяться, пока не приблизится к равновесному состоянию, охарактеризованному в Fn поколении. Каким закономе рностям это состояние отвечает?
Ю. А. Филипченко в 1924 г. показал, что в дигенной панмиктической популяции для достижения состояния равновесия требует ся установление конкордантного (согласованного) соотношения ча стот гамет. При конкордантном соотношении выдерживается раве нство g11 X g33 поколении показатель дискордантности d отлича ется от нуля и составляет —0,02 (табл. 3.11). В каждом последу ющем поколении этот показатель уменьшается в 2 раза. Практичес ки достаточно 6—7 поколений случайных скрещиваний, чтобы он приблизился к нулю. Но такая скорость будет иметь место при отсутствии сцепления между анализируемыми генами. Если же эти гены сцеплены, то чем больше сила сцепления, тем медленнее дигенная панмиктическая популяция будет приближаться к состоя нию равновесия (рис. 3.10).
Дискордантное соотношение частот гамет, измеряемое величи ной d, может вызываться не только сцеплением разных генов, но их неслучайной ассоциацией в результате действия отбора или других факторов. Это явление принято называть неравновесием по сцепле нию или гаметическим неравнове сием. Наличие гаметического не равновесия обусловливает корре ляцию между частотами аллелей разных генов в популяции. При этом сдвиг в частотах аллелей од ного гена будет приводить к изме нению частот аллелей другого ге на, с ним ассоциированного. Вза имозависимость популяционногенетических процессов, протека ющих по разным локусам, в силу
неравновесия |
по |
сцеплению |
|
Л. А. Животовский |
определяет |
||
как гаметическую интеграцию. |
|||
Для |
нахождения |
равновесных |
|
частот |
генотипических классов |
||
в панмиктической популяции при дигенных различиях нужно произ вести перемножение трехчленов
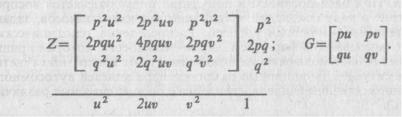
(биномиальных |
распределений, |
по |
Харди—Вайнбергу): |
(p2+2pq+q2) (u2+2uv+v2). Для определения |
равновесных гамети- |
||
ческих частот следует перемножить двучлены: (p+q) (u+v). Тогда получим следующие матрицы генотипических и гаметических частот:
Легко видеть, что в состоянии равновесия принцип конкордантности частот гамет выражается тождеством рu * qv=pv * qu.
При полном доминировании в дигенной панмиктической попу ляции будут образовываться 4 фенотипических класса особей. Их частоты найдем при суммировании частот соответствующих гено типических классов:
A-B-:(1-q2)(1-v2); |
А-bb: |
(l-q2)v2; ааВ-: q2(l-v2); |
|
ааbb: q2v2. |
|
Можно путем дальнейшего суммирования показать, что частоты четырех фенотипических классов по каждому из двух пар альтер нативных признаков в отдельности составляют:
A-: 1-q2; аа: q2; B-: 1-v2; bb:v2.
У кур розовидная форма гребня (А) доминирует над листовид ной (а); наличие хохла (В) доминирует над его отсутствием (b). В одной из панмиктических популяций обнаружены особи 4 феноти пических классов в следующих количествах (А. С. Серебровский, 1970):
A-B- |
А-bb |
ааВ- |
ааbb |
130 |
277 |
119 |
232 |
При условии панмиксии и отсутствии неравновесия по сцепле нию между аллелями генов А и В теоретически ожидаемые числен ности особей четырех указанных классов должны составить:
133,5 274,0 114,8 235,7.
При этом частоты аллелей равны: P(a) = 0,32; q(a) = 0,68; u(b)=0,18, v(b) = 0,82; гаметические частоты: g11 = 0,06; = 0,26; g3l =0,12; g33 = 0,56. Наблюдаемое распределение особей не отличается стати стически достоверно от теоретически ожидаемого (х2 = 0,33; d f = 4 - 2 - 1 = 1;p>0,05).
! В данной главе были рассмотрены закономерности наследова ния в панмиктической популяции при моногенных и дигенных раз личиях. Под наследованием в популяции подразумевается воспро изведение в ряду поколений частот генотипических классов, задан ных данными частотами аллелей. Распределения генотипических частот описываются формулой Харди—Вайнберга, которая прини мает различные модификации в зависимости от конкретных генети ческих ситуаций (моногенные различия: пара аллелей аутосомного гена, множественные аллели, сцепление с полом; дигенные различия и проч.).
Были описаны отклонения эмпирически найденных частот гено типических классов от теоретически ожидаемых по Харди—Вайн бергу (случаи окраски шерсти у кошек Лондона, окраски яиц в по пуляции кайры). Причины отклонений заключались в дифференци альном действии отбора, избирательной гибели особей отдельных генотипических классов, чаще всего рецессивных гомозигот.
Глава 4 ИЗМЕНЕНИЕ ЧАСТОТ АЛЛЕЛЕЙ
ВПОПУЛЯЦИИ
4.1.ФАКТОРЫ МИКРОЭВОЛЮЦИИ
!В качестве эволюционных факторов, способных изменять генотипический состав популяций, выступают: мутационный процесс; «волны жизни» — колебания численности особей в популяциях; изоляция — возникновение барьеров внутри и между популяциями; наконец, естественный отбор, который играет роль направляющего фактора эволюции. Элементарным эволюционным явлением следу ет считать длительное, векторизованное преобразование генотипического состава популяции.
Изменение частот аллелей в популяции происходит под непо средственным воздействием мутационного давления, отбора, гене тического дрейфа и процесса миграции. В данной главе будут рассмотрены количественные закономерности действия этих фак торов в панмиктической популяции при моногенных различиях. Именно со сдвига частот аллелей отдельных генов начинается изменение генетической структуры популяции.
4.2. МУТАЦИОННОЕ ДАВЛЕНИЕ
Скорость, или частота мутирования, измеряется долей гамет на поколение, в которых произошли мутационные изменения данного гена.
Различают скорости прямого и возвратного мутирования. Их обозначают греческими буквами m и v: А <->а. Пусть m=1 * 10-5 ,а
-6 V
V = 1* 10 . Это означает, что на миллион гамет будет приходиться в среднем 10 с вновь возникшими рецессивными и 1 с вновь возник шей доминантной мутациями анализируемого гена.
Примем частоту доминантной аллели в Fn поколении за рп. Тогда прирост частоты рецессивной аллели за счет прямого мути рования в следующем Fn+1 поколении составит mрп. Однако убыли частоты доминантной аллели на эту величину будет противостоять

ее прирост за счет возвратного мутирования vqn. В результате разница в частотах аллелей между двумя смежными поколениями выразится так: q=mpH—vqH. Нетрудно видеть, что такая ситуация приведет в конечном счете к равновесию, при котором убыль в частотах аллелей будет покрываться их прибылью, т. e . q = 0 и mр„= =vq„. В состоянии равновесия p/q=v/m; q=m/(m+v); р=v/(v+m).
Пусть |
значения m и v будут такие, как указано выше: 10-5 |
и 1O-6. |
Тогда равновесные частоты аллелей составят p=0,09; |
q=0,91. Графически эта ситуация отражена на рис. 4.1. Из графика следует, что q положительна при q<q' и отрицательна при q>q.
Необходимо иметь в виду, что истинные обратные мутации, ; т. е. мутации в том же гене, в котором произошла прямая мутация, составляют лишь малую часть от общего числа. Абсолютное боль шинство реверсий обусловлено мутациями других генов, дающих супрессорный эффект.
! Итак, мутационное давление оказывает крайне слабое влияние на изменение частот аллелей в популяции в силу низких скоростей процесса спонтанного мутирования. Картина, правда, может стать иной при возникновении нестабильности генетического материала (см. разд. 8.3 и 8.4). Эволюционное значение мутационного процесса заключается, однако, не в преобразовании аллельных частот, а в со-|
здании разнообразия аллелей и появлении новых генов. Мутационный процесс — поставщик материала для действия отбора, и в этом качестве он выступает как фактор эволюции.
43. ДЕЙСТВИЕ ОТБОРА
Эффективность действии отбора при полной элиминации рецессив ных гомозигот. Полная элиминация рецессивных гомозигот, т. е. их устранение от оставления потомства, может иметь место при нали чии рецессивных летальных мутаций или мутаций стерильности.! Элиминацию можно осуществлять и искусственным путем.

Обратимся к примеру, который уже был рассмотрен в гл. 3. Речь идет о популяции крупного рогатого скота породы Шортгорн. различия в окраске шерсти между животными трех генотипических классов контролируются парой аллелей, частоты которых на протя жении 30 лет оставались близкими к значениям: р=0,1; q=0,3. Особи с белой окраской характеризуются, как было отмечено, сниженной фертильностью и некоторыми другими нежелательными свойствами. Представим, что селекционеры пришли к решению избавиться в породе Шортгорн от животных данного класса. По скольку имеет место неполное доминирование, задачу можно было решить радикальным способом, устранив от оставления потомства не только гомозиготных, белых производителей, но и гетерозигот ных, чалых родительских особей. Тогда все представители породы Шортгорн приобрели бы одинаковую, по выражению животново дов, «рубашку» — красного цвета. Однако такой путь привел бы к недопустимым убыткам. Поэтому рациональнее выбраковывать в каждом поколении из племенного стада бычков и телочек только белой масти. Но прежде необходимо получить точный прогноз, в какой пропорции будут появляться телята трех генотипических классов, спустя, скажем, 10, 20 или 50 поколений с начала отбора.
Решим задачу сначала в общем виде. При этом будем иметь в виду, что оставляемые на племя производители красной и чалой масти скрещиваются случайно, т. е. популяция, несмотря на отбор, по-прежнему поддерживается панмиктически. Элиминация рецес сивных гомозигот приводит каждый раз к изменению частот оста ющихся двух генотипических классов. Ход решения задачи дан в табл. 4.1. В этой таблице генотипическая структура популяции представлена в каждом поколении на двух этапах — до и после выбраковки рецессивных гомозиготl . Частотыl аллелей устанавлива ются по формулам p = P+ /2H; q = Q + /2H. Соотношение трех ге нотипических классов на этапе до выбраковки соответствует фор муле Харди—Вайнберга.