

3 курс / Топографическая анатомия и оперативная хирургия / Хирургическая_анатомия_венечных_артерий
.pdfВ литературе мы не встрети ли материалов, раскрывающих причину отхождения ВА от лице вых синусов аорты. Между тем A. Gittenberger-de Groot и соавт. (1983) утверждают, что анализ данных литературы позволяет с уверенностью констатировать постоянство отхождения ВА от лицевых синусов аорты.
Анализ данных литературы показал, что до настоящего вре мени не разработано приемлемой (неуязвимой) теории, позволяю щей объяснить соединение второ
го и третьего компонентов венечно-артериальной системы (и в норме, и при аномалиях), что, по-видимому, обусловлено: 1) попытками объяснить этот процесс изолированно от септации выводных отделов, артериальных стволов и отверстий либо механистичес ки - без понимания сложности данного процесса; 2) отсутствием единства во взглядах на процесс сепарации артериального ствола и выводных отделов и терминологическими разногласиями относительно структур указанного отдела сердца.
Перечисленные процессы протекают не сами по себе, а в тесном взаимодействии. Поэтому понятно, что изолированное толкование указанных процессов септации, се парации, интеграции и других является основной причиной разногласий. Отсюда не обыкновенная, на наш взгляд, прозорливость R. Sahner, предсказавшего, что «сердце необычайно сложно, чтобы приспособиться ко всем нашим теориям», разрабатывае мым в отрыве от сопредельных процессов.
5. Соединение системного и коронарного кругов кровообращения. Септация артериальных отверстий и выводных отделов желудочков сердца
Механизм присоединения венечно-артериального сплетения к аорте и процессы септации и сепарации артериальных стволов, артериальных отверстий и выводных отделов тесно взаимосвязаны и взаимообусловлены. Разработкой этого вопроса за нимались исследователи из Лейденского университета (А. Гиттенбергер-де Хруут, 1990; M.Bartelings et al., 1988; A. Bogers et al., 1988; A. Bogers, 1989). Хотя до сих пор многие аспекты соединения системного и коронарного кругов кровообращения остаются неясными, авторам удалось субстанционально опровергнуть бытовавшие ранее предположения.
Описание процессов развития конотрункуса связано с немалыми трудностями. До недавнего времени большинство исследователей (Г.Анселъми и др., 1981; Ю.С.Петросян и др., 1982; В. И. Бураковский и др., 1987; Р. Ван Прааг, 1990; Д. Э. Фелдмане и др., 1990; R. Collett and J. Edwards, 1949; L. Van Mierop, 1963, 1978; R. Van Praagh and S. Van Praagh, 1965; D. Gooret al., 1972; R.Anderson et al., 1974; G. Rosenquist et al, 1976; L.Kutsche and L. Van Mierop, 1987) полагали, что септация конотрункуса осуществля ется на трех уровнях: артериального ствола, артериального отверстия и конотрункуса.
Между тем из-за терминологических и структурных расхождений в толковании развития этого региона (конотрункуса) даже среди эмбриологов однозначно понять,
что образуется и как |
идет этот процесс не представляется возможным (T.Pexieder |
et al., 1989). Поэтому |
в дальнейших рассуждениях мы опирались на определения |
Н. Laane (1978) и М. Bartelings и соавт. (1988), которые предпочли ориентироваться
29

не на гипотетические границы между сегментами сердца, а на гистологически четко определяемую границу между миокардиальным конусом (выводным отделом) и мезенхимальным артериальным стволом (трункусом).
М. Bartelings и соавт. (1986, 1988, 1989) показали, что процесс разделения артери альных стволов, отверстий и выводных отделов един и осуществляется не путем септации отдельных сегментов, а за счет сепарации их одной структурой - аортолегочной перегородкой. Согласно материалам этих авторов, первоначально (на стадии развития 6-7-миллиметрового эмбриона) артериальный ствол и конус располагаются на разных уровнях (рис. 6,А). Первый из них имеет мезенхимальную стенку, второй - миокардиальную. Уровень артериального отверстия следует констатировать между указан ными сегментами и тканями, образующими эти сегменты. Как следует из рис. 6, А, на данном этапе развития сердца артериальное отверстие располагается в одной плоскос ти. По нашим предположениям, приведенным выше, в той же плоскости располагает ся и перитрункальное кольцо (рис. 6,А; 7,А).
В норме, по мере роста сердца, миокардиальный край вытягивается вверх в ви де двух выступов, вследствие чего его верхний край деформируется, приобретая седловидную форму. Граница миокардиального края вентрально достигает почти
Рис. 6. Сепарация артериальных отвер стий по М. Bartelings (1990). Схематичес кое изображение аортального мешка, вы водной камеры (ВК) и желудочково-арте- риального соединения на разных этапах формирования сердца (вид сзади) (предсердный сегмент удален).
А - потенциальные уровни сепарации: I - артериаль ные стволы, II - артериальные отверстия и клапаны, III - ВК. Б - изменение пространственных взаимоот ношений на уровне артериальных отверстий обу словливает септацию аортолегочной перегородкой на уровне, показанном стрелкой с пунктиром. В ре зультате такого разделения образуются очень корот кий легочный ствол, отдающий правую и левую ле гочные артерии (ЛА) и 6-е аортальные дуги сразу, и относительно более длинная восходящая аорта.
3, 4 и 6 - третья, четвертая и шестая аортальные ду ги, ПК - приточная камера, АВО - общее атриовентрикулярное отверстие.
30

Артериальное
отверстие
Рис. 7. Взаимоотношения меж ду перитрункальным кольцом и артериальными отверстиями до
(А) и после (Б) их сепарации.
А— клапан аорты, ЛА — клапан легоч ной артерии, 1 и 2 - лицевые синусы аорты. Треугольными стрелками огра ничены правый (светлая) и левый (тем ная) края ретробульбарного сегмента вокругпредсердного венечно-артериально- го кольца. X - перерыв бульбовентрикулярного кольца, образующего в дефини тивном сердце ПМЖВ.
Звездочками ограничен фрагмент перитрункального кольца, опускающийся (показано светлой изогнутой стрелкой) ниже артериального клапана. Толстой светлой стрелкой показана приподня тость переднего края легочного клапана (выше исходной позиции, показанной пунктиром).
уровня четвертых аортальных дуг, а дорсально - шестых. От этих наиболее высту пающих точек граница миокардиального края тянется направо и налево вокруг просвета артериального ствола (см. рис. 6,Б).
Вследствие этого изменяется и конфигурация артериального отверстия: аорталь ная часть отверстия наклоняется относительно легочной. В связи с этим изменяется конфигурация и перитрункального кольца. Впоследствии такое изогнутое взаимоот ношение будущих сепаратных отверстий общего артериального отверстия дополняет ся их перекрутом в обратных направлениях, в результате чего плоскость будущего аортального клапана ориентирована кзади и вправо, а клапана легочного ствола - кпе реди и влево (рис. 7,Б; 8).
Разделение артериальных отверстий начинается вслед за такой деформацией еще неразделенного общего артериального отверстия и осуществляется экстракардиальной конденсированной мезенхимой, которая перекидывается в виде мостика между двумя миокардиальными выступами, образуя при этом арку над инфундибулярными отделами правого и левого желудочков. Поскольку данная структура ориентирована косо, а конфигурация верхнего края миокардиального конуса седловидно деформи-
Рис. 8. Пространственные взаимоотношения между аортальным (А) и легочным (Р) кла панами, воспроизведенные от эмбриона че ловека длиной 9,5 мм методом графической реконструкции (по М. Bartelings, 1990).
Легочный клапан расположен более или менее попереч но, краниально и кпереди-влево от аортального, распо лагающегося ближе к фронто-сагиттальной проекции и продолжающегося каудально и кзади-вправо.
KVOT - выводной отдел правого желудочка; арка, пред ставляющая аортолегочную перегородку (показана звездочкой), делит артериальные отверстия (показаны стрелками) и посылает свои ножки (обозначены крести ками) в выводной отдел. RA - правое предсердие, LA - левое предсердие, РА - легочные артерии, AD - дор
сальные аорты, 4R , 4L , и 6L — 4-я правая, 4-я левая и 6-я левая аортальные дуги.
31
рована, она разделяет выводной отдел (подлегочный инфундибулум) правого желу дочка и аортальный мешок (восходящую аорту). Ввиду такого ориентированного раз деления к моменту сепарации легочный ствол практически не имеет длины, как и подаортальный инфундибулум, и, наоборот, восходящая аорта и подлегочный инфунди булум имеют достаточную протяженность (см. рис. 6,Б). Иными словами, согласно М. Bartelings и соавт. (1989), количественные взаимоотношения и ориентационные особенности восходящей аорты и легочного ствола, артериальных клапанов и вывод ных отделов, определяющиеся к моменту сепарации, практически идентичны тако вым в зрелом сердце.
Чтобы объяснить механизм соединения венечно-артериального сплетения с разде ленными артериальными стволами, представляется уместным коротко упомянуть о сроках развития венечно-артериального субэпикардиального сплетения.
Долгие годы предполагали, что процесс присоединения осуществляется до сепара ции артериальных стволов (J. Goldsmith and H. Buttler, 1937; S. Viragh and E. Challice, 1981). G. Hutchins и соавт. (1988) и A. Bogers (1989) показали, что полностью сфор мированное перитрункальное кольцо обнаруживается только после сепарации артери альных отверстий. Эти данные опровергают возможность присоединения ВА к артери альным стволам до их разделения8 . Отсюда это соединение происходит только после разделения артериальных отверстий. И тут, мы полагаем, важно подчеркнуть, что процесс соединения ВА с артериальными стволами определяется не столько процес сом сепарации, сколько детерминирован изгибом и ориентацией плоскостей аорталь ного и легочного клапанов.
Как следует из рис. 6, воспроизводящего пространственные взаимоотношения ар териальных отверстий, перитрункального кольца, сегментов сердечной трубки и тка ней, образующих их стенки, эти взаимоотношения на данном этапе значительно изме нены (по сравнение с предыдущими этапами) и достаточно сложны: 1) вследствие изме нения ориентации артериального отверстия (и, соответственно, ориентации разделен ных артериальных отверстий) меняется и отношение к нему перитрункального кольца; 2) ввиду изменения плоскостей, определяющих границы тканей, образующих стенки различных сегментов конотрункуса, изменяется отношение перитрункального кольца и к этим стенкам. Как уже отмечалось, первоначально перитрункальное кольцо распо лагается в одной плоскости с границей между конусом и трункусом. После изменения конфигурации отверстия, образуемого миокардиальной тканью, его верхняя граница в легочной части трункуса проходит выше перитрункального кольца и прикрывает собой эту часть. В результате такого перемещения тканей этот отдел приобретает внутрен нюю (мезенхимальную) и наружную (миокардиальную) стенки. Пенетрирование арте риального ствола в этой (ставшей двухслойной) части стенки маловероятно.
С другой стороны, в результате изгиба неразделенного артериального отверстия перитрункальное кольцо начинает тесно прилегать к той части артериального ствола, из которой в будущем образуется восходящая аорта. Область такого прилежания пе ритрункального кольца - это зона непосредственно над верхним краем мышечной гра ницы выводного отдела, соответствующая в будущем области синотубулярного соеди нения, чуть выше уровня образования лицевых заслонок аортального клапана9 .
Вероятность пенетрации эластичной (однослойной) стенки аорты перитрункальным кольцом в этой области весьма возможна1 0 .
8Этот факт делает понятным происхождение редких аномалий ВА, заключающихся в отхождении ЛВА от первого лицевого синуса аорты или ПВА - от второго и их последующем следовании между аортой и ЛС. Формирование подобных аномалий, по-видимому, обусловлено аномальным присоединением пери трункального кольца к несоответствующему синусу в результате перекидывания кольца через аортолегочную перегородку (после разделения артериальных клапанов).
9В области нелицевого синуса аорты свободный сегмент перитрункального кольца не прилежит тесно к аортальной части артериального ствола. Кроме того, их плоскости в этой области не совпадают.
1 0 При более высоком расположении кольца возможна эктопическая пенетрация стенки аорты.
32
Таким образом, процесс соединения венечно-артериального сплетения с одним из ар териальных стволов детерминирован изгибом неразделенного артериального отверстия и ориентацией плоскостей разделенных аортального и легочного клапанов. Эти ориентационные особенности, на наш взгляд, являются основой сближения перитрункального кольца с восходящей аортой и ее лицевыми синусами (см. рис. 7,Б). Избирательная пенетрация восходящей аорты обусловлена тем, что на данном этапе развития легочный компонент артериального ствола прикрыт миокардиальными выступами (см. рис. 6,Б).
Мы полагаем, и ниже это будет показано, что в основе закладки аномального отхождения левой ВА от ЛС лежат изменения пространственных взаимоотношений аор тального и легочного компонентов делящегося артериального отверстия и изменения взаимоотношений перитрункального кольца с этими структурами.
Таким образом, избирательное присоединение перитрункального кольца к тому или иному артериальному стволу (в норме - к аорте) обусловливается интимным прилежани ем указанных структур, а также строением (числом слоев) стенок артериальных стволов.
Специфичное (избирательное) присоединение перитрункального кольца к области формирования лицевых синусов аорты в норме обусловлено седловидной конфигураци ей артериального отверстия и перекрутом плоскостей разделенных артериальных от верстий, а также конфигурацией миокардиального края конуса, окружающего эти от верстия, определяющих тесное прилежание этих структур (лицевых синусов аорты и перитрункального кольца).
Уровень отхождения ВА (и расположения устьев ВА) в норме обусловлен специфи ческой конфигурацией верхнего края миокарда (окружающего артериальные стволы в области желудочково-артериального соединения), определяющей пространственные взаимоотношения перитрункального кольца и артериальных стволов и тесного его при лежания к области синотубулярного соединения аорты.
Время формирования венечно-артериального соединения зависит от последова тельности изгиба артериального отверстия, его сепарации, формирования перитрун кального кольца и пенетрации стенки аорты.
6. Формирование дефинитивной системы венечных артерий
Как известно, дефинитивное строение ВА предполагает наличие двух (правой и ле
вой) ВА, отходящих от лицевых синусов |
аорты (Н'. А. Джавахишвили |
и М.Э.Кома- |
|||||
хидзе, 1967; Д.Лужа, 1973; |
В.В.Кованое и |
Т.Н.Аникина, 1974; |
И.Х.Рабкин, |
1977; |
|||
T.James, |
1961; |
Z.Vlodaver et |
al., 1975; W.McAlpine, 1976; M.Trivellato |
et al. |
1980; |
||
H.Neufeld |
and |
A. Schneeweiss, |
1983; P.Angelini, 1989; A.Bogers, |
1989). |
Какова же |
||
судьба венечно-артериального сплетения? Каковы механизмы и пути его трансформа ции в дефинитивную систему ВА? Несмотря на то, что фрагментарно эти вопросы ос вещены в единичных сообщениях (S. Van Praagh et al., 1982; P.Angelini, 1989; A. Bogers, 1989), полного ответа на указанные вопросы в литературе нет, как нет и ма териалов, освещающих параллельное развитие желудочкового комплекса и формиро вание ВА, и данных об одновременной оценке развития ВА и процессов развития конотрункуса. Единственное исключение, пожалуй, составляет комплексное исследова ние ученых из Лейденского университета, освещающих эти вопросы параллельно
(A. Gittenberger-de Groot et al., 1987; Bartelings et al., 1988; A. Bogers, 1989). Тем не ме нее, при несомненном успехе, достигнутом этими исследователями, им также не уда лось исчерпывающе расшифровать код хитросплетений развития этих независимых и одновременно сопряженных и тесно переплетающихся процессов.
Для объяснения основных механизмов и путей формирования дефинитивной сис темы ВА мы проанализировали этот процесс параллельно с формированием желудоч кового комплекса и далее в сопоставлении с большой группой сердец с полным разво-
33

ротом оси конотрункуса с обеими сторонами петлеобразования сердца. Как уже отме чалось, по мере завершения процесса петлеобразования сердца образуется общее ве- нечно-артериальное сплетение, обособленное от системного кровообращения. Схема тически сплетение представляет собой ряд колец, сливающихся в области бульбовентрикулярной складки (см. рис. 2, 3). Принципиально важно различать пять базовых колец: вокругартериальное (перитрункальное) (Ах ), вокругбульбарные (А2, А3 ), бульбовентрикулярное (межжелудочковое, В), вокругжелудочковые (D и Е) и вокругпредсердное (предсердно-желудочковое, С) (см. рис. 3).
До недавнего времени процесс образования ВА интерпретировали как появление вы ростов аорты, которые в дальнейшем присоединяются к сосудистому сплетению. Соглас но материалам, представленным A. Bogers (1989), процесс соединения венечно-артери- ального сплетения с аортой предусматривает прорастание проксимальных ВА в аорту из вне (см. рис. 5). Иными словами, не венечные артерии растут из аорты, как это предпо лагали все предыдущие авторы, а перитрункальное кольцо индуцирует проксимальные ВА, прорастающие в стенку аорты снаружи вовнутрь. Мы полагаем, что присоединение ВА к аорте осуществляется за счет пенетрирования ее стенки самим вокругартериальным (перитрункальным) кольцом (см. рис. 7, Б), без участия дополнительных прокси мальных ВА. Поскольку кольца-фрагменты представляют часть единого венечно-арте- риального сплетения, процесс интеграции перитрункального кольца (интеграционное кольцо) в системную циркуляцию одновременно вовлекает в артериальную циркуляцию все венечно-артериальное сплетение (рис. 9, 10). При этом остальные кольца трансфор мируются в коммуникационные фрагменты (они начинают нести коммуникационную функцию для осуществления венечного кровотока от аорты к миокарду и для осуществ ления связи друг с другом). На хронологии присоединения ВА к аорте детальнее мы ос тановимся ниже. Здесь же попытаемся описать дифференциацию сплетения по мере осу ществления преобразований в сердце, поскольку соблюдение последовательности этих изменений продиктовано неоднозначностью изменений в первичном сплетении.
Рис. 9. Формирование дефинитивной системы венечных артерий после инкорпорации синус ной части в состав правого желудочка и завершения роста выводных отделов желудочков.
По мере интегрирования синусной части в состав ПЖ правожелудочковый фрагмент бульбовентрикулярного кольца
(В) и вокругбульбарные кольца разобщаются.
По мере роста выводного отдела правого желудочка вокругбульбарные кольца (А2, А3 ) разрываются. По мере роста ле вого желудочка происходит разрыв вокругжелудочковых колец (D, С). А - вид спереди, Б - вид сзади, В - вид сбоку, справа. T - трехстворчатый клапан, М - митральный клапан; Р - легочная артерия, А - аорта.
Первые изменения в сплетении касаются бульбовентрикулярных и вокругжелу дочковых колец. По мере образования синусной части правого желудочка (ПЖ) и об разования раздельных (правого и левого) атриовентрикулярных отверстий бульбовентрикулярные круги сплетения растягиваются, постепенно рассасываются и разрыва ются (рис. 11). Разорванные концы в дефинитивном сердце формируют конкретные ВА, а сохранившаяся часть сливается с вокругпредсердным кругом (рис. 12, круг С).
34

Рис. 10. Различия в строении венечно-артериальной системы в зависимости от стороны рота ции конуса (от исходной позиции).
А. При ротации конуса по часовой стрелке (в нормальном сердце, например) правая венечная артерия (ПВА) отходит от 1-го лицевого синуса, а левая (ЛВА) - от 2-го.
Б. При ротации конуса против часовой стрелки (как это бывает в сердцах с транспозицией аорты и легочной артерии, например) ПВА отходит от 2-го лицевого синуса, а ЛВА — от 1-го.
Фрагменты а и б демонстрируют варианты отхождения конусной артерии (КА) от ПВА или от 1-го лицевого синуса само стоятельно (в нормальных сердцах) и варианты отхождения ОВ (от ЛВА или ПВА) в сердцах с транспозицией аорты и ле гочной артерии. Объяснение механизма формирования указанных типов соединения венечных артерий см. в тексте.
ОВ - огибающая ветвь, ДВ - диагональная ветвь, ПМЖВ и ЗМЖВ - передняя и задняя межжелудочковые ветви.
Процесс интеграции правожелудочковой порции синусной части примитивного желу дочка (приточного компонента сердечной трубки) в состав ПЖ в литературе не имеет однозначного толкования. Здесь мы ограничимся описанием собственных представле ний об этом процессе, основанном на сопоставлении сердец с атрезией правого атриовентрикулярного отверстия (АПАВО), единственным желудочком (ЕЖ) левожелудочкового типа и нормальных сердец (Н). Согласно нашим представлениям, сердца ука занных групп отличаются по формированию желудочков или, точнее, представляют этапы формирования желудочков (рис. 13). В сердцах с АПАВО рост единственного (левого) желудочка осуществляется равномерно эксцентрично по всей окружности левожелудочкового контура. В норме же имеет место эксцентрический рост строго опре-
Рис. 11. Изменения в венеч- но-артериальном сплетении по мере образования синус ной части правого желудочка и образования раздельных (правого и левого) атриовентрикулярных отверстий.
Объяснение в тексте.
35

Рис. 12. Формирование дефинитивных венечных артерий.
Объяснение в тексте.
деленной зоны - небольшого сопредельного сегмента стенки желудочка в области сты ка ткани примитивного желудочка и бульбуса под и над правой частью общего атриовентрикулярного отверстия, в результате чего этот бурно растущий регион в сердцах с ЕЖ образует соответствующую область роста в составе примитивного желудочка (зона роста находится у стыка, но не распространяется на сам стык). В норме же зона роста расположена в области стыка бульбуса и примитивного желудочка. Поэтому рост этой
зоны обусловливает перемещение порции синусной |
части приточного компонента |
||
в бульбус, раскрывает бульбовентрикулярное |
окно |
и перемещает |
бульбус вперед, |
то есть ротирует его против часовой стрелки |
(И. И. Беришвили и др., |
1991) (рис. 14). |
|
Указанное предположение во многом основано на сопоставлении изменений в первич ных кольцах венечно-артериального сплетения в сердцах с АПАВО, ЕЖ и в норме (Н).
Всердцах с АПАВО (исходный этап) вокругартериальный (Ах), вокругбульбарные (А2,
А3 ), бульбовентрикулярный (В) и вокругжелудочковые (D и Е) круги сплетения схо
дятся в ретробульбарной области. В сердцах же с ЕЖ и Н нормальные взаимоотноше ния с вокругпредсердным кругом (С) сохраняют только вокругартериальные и вокруг бульбарные круги. По мере эксцентрического роста приточной части ПЖ правая сто рона бульбовентрикулярного круга (В) теряет связь с указанными кругами. Ввиду пе реднего перемещения (ротации) бульбуса от исходной позиции резко удаляется пра вый край вокругбульбарного и вокругартериального кругов (А). При этом они, как от мечалось, не теряют связи с вокругпредсердным кругом. От них удаляется правый край бульбовентрикулярного круга (В), в результате чего он уже не сливается с круга ми группы А (см. рис. 13, 14). Еще больше удаляется от этих кругов правый край вокругжелудочковых кругов (D и Е). Эти данные, кроме изменения в строении бульбо вентрикулярного и вокругбульбарных кругов, свидетельствуют о том, что формирова ние дефинитивного ПЖ осуществляется не перемещением правой части общего атрио-
36
Бульбовентрикуляр- |
Атрезия правого атрио- |
Единственный желудочек |
Норма |
ная петля |
вентрикулярного отверстия |
|
|
Рис. 13. Изменения в строении венечно-артериальной системы на различных этапах формиро вания желудочков (на примере сердец с различным строением желудочков).
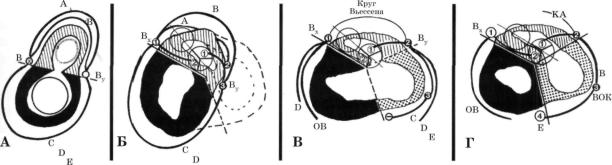
А - срез сердца (тотчас ниже атриовентрикулярных клапанов) на этапе образования бульбовентрикулярной петли; Б - срез сердца с атрезией правого атриовентрикулярного отверстия; В - срез сердца с единственным желудочком левожелудочкового типа; Г - срез нормального зрелого сердца. Вокругбульбарные (А), бульбовентрикулярные (В) и во кругжелудочковые (С, D, Е) венечно-артериальные кольца соединяются в точках Вх и By на линии стыка примитив ных бульбуса и желудочка. Линия Вх-Ву - ретробульбарный сегмент сплетения. Во всех сердцах сохраняется соеди нение остатков этих колец в точке Вх (в точке 1). Во всех остальных сердцах ретробульбарный сегмент венечно-арте- риального сегмента разорван (адсорбирован) задним артериальным клапаном (на рисунке - аортой). Его остатки пред ставлены левой ВА (простирающейся между 2-м лицевым синусом аорты и точкой 1) и правой ВА (простирающейся между 1-м лицевым синусом и точкой 2).
Начиная с сердец с атрезией правого атриовентрикулярного отверстия (Б) бульбовентрикулярное кольцо уже не соеди няется с остальными кругами в точке 2. Оно соединяется (точка 3) только с вокругпредсердным кольцом. По мере фор мирования синусной части (точечная штриховка) рвутся вокругжелудочковые круги и удаляются друг от друга точки 2 и 3. Разрыв круга Вьессена во многом определяется размерами выпускной камеры (В). Г - окончательное состояние системы венечных артерий в дефинитивном сердце. Разорванные вокругжелудочковые круги образуют дефинитивные ВА. При этом ОВ, ДВ и ВТК, являющиеся дериватами кругов С, D и Е, отходят от точки 1.
Всвязи с серьезными структурными изменениями в строении правого желудочка ВА, питающие его стенки, отходят от разных точек: конусная артерия (КА) - от точки 2, ветвь острого края (ВОК) - от точки 3 и ЗМЖВ - от точки 4 в об ласти задней межжелудочковой борозды. Из приведенной схемы понятно, что ЗМЖВ обнаруживается в нормальных сердцах и в некоторых сердцах с единственным желудочком, но не выявляется при атрезии правого атриовентрику лярного отверстия. В этих сердцах артерия, располагающаяся на границе желудочка и выпускника справа-сзади, представляет собой ветвь острого края (отходящую от точки 3).
Всвязи с развитием инфундибулярной камеры круг Вьессена в нормальном сердце, как правило, разорван.
вентрикулярного канала в сторону ПЖ, а благодаря нескольким факторам: экстенсив ному росту ограниченного участка приточного компонента, переднему перемещению задней стенки бульбуса и его росту, а также раскрытию бульбовентрикулярного отвер стия вследствие перечисленных выше изменений (см. рис. 14). Приведенная состав ная форма роста ПЖ также отличается от общепринятой (только ростом бульбуса), приведенной S. Rao (1982). Если зона экстенсивного роста располагается полностью в составе примитивного желудочка, процесс развития желудочкового миокарда осуще ствляется без интеграции тканей приточного компонента в состав ПЖ, что в итоге обусловливает формирование сердец с единственным желудочком.
Таким образом, процесс формирования ПЖ влечет за собой разрыв вокругжелудочковых кругов, осуществляя тем самым формирование задней межжелудочковой ветви (ЗМЖВ) правой ВА (из правого компонента круга Е) и диагональной ветви левой ВА (из левого компонента круга Е), а также заднебоковой ветви правой ВА (из право го компонента круга D) и ветви тупого края левой В А (из левого компонента круга D). В результате перемещения правого края вокругбульбарного круга из него формирует ся ветвь острого края правой ВА (см. рис. 9 и 13). Левый край этого круга формирует переднюю межжелудочковую ветвь (ПМЖВ) левой ВА, теряющей связь с ветвью ост рого края только по мере роста трабекулярной части и всего ПЖ .
Как следует из приведенных материалов, процесс роста желудочков и связанный с ним разрыв вокругжелудочковых кругов ответственны не только за формирование де финитивных ВА, определяемых в зрелом сердце, но и участвуют в формировании из вестных (правого, левого и сбалансированного) типов (доминантности) кровоснабже ния миокарда. Этот же процесс определяет и то, ветвью какой (правой или левой) ВА
37

Рис. 14. Схема перемещения точек соединения венечно-артериальных колец с ростом правого желудочка.
А - известная схема роста желудочков из книги S. Rao «Tricuspid atresia» (N. Y., 1982, P. 32). Схема предусматрива ет концентрический рост желудочковых полостей в течение внутриутробной жизни. Как показали сопоставления сер дец с различной степенью дифференциации желудочкового комплекса (см. рис. 13), формирование правого желудоч ка отличается от такового, приведенного в книге. Для его формирования необходимо развитие синусной части и инте грация последнего в состав правого желудочка. За счет интенсивного роста этой части правого желудочка формирует ся приточный отдел и смещается вперед выводной. Этот же процесс предполагает пространственную переориентацию
ираскрытие бульбовентрикулярного окна. Будучи первоначально отверстием между бульбусом и примитивным желу дочком, в дефинитивном сердце оно разграничивает приточный и выводной отделы. Б - общая схема формирования и роста желудочкового комплекса, разработанная нами на основании приведенных данных. В соответствии с приведен ными материалами нами проанализированы изменения в венечных артериях. В - раздельная схема роста приточного
ивыводного отделов правого и левого желудочков.
Первоначально вокругбульбарные (А), бульбовентрикулярное (В) и вокругжелудочковое (Е) кольца сливаются (см. в бульбусе (Б), из которого формируется выводной отдел нормального сердца).
Перемещения точки отхождения вокругбульбарного кольца (А) с ростом ПЖ определяют точки Ах -А5 . А - точка отхождения конусной артерии от правой ВА. Перемещения точки отхождения бульбовентрикулярного кольца (В) с рос том приточного отдела правого желудочка определяют точки Bj—В5. В — точка, отхождения ветви острого края. Е — точ ка отхождения ЗМЖВ.
Формирование дефинитивного ЛЖ из примитивного желудочка осуществляется за счет концентрического роста послед него (при утере правожелудочковой части синусного отдела и приобретении аортальной части артериального конуса).
На схеме Б: 0Y - проекция задней межжелудочковой перегородки, ОХ - проекция передней межжелудочковой перегород ки, 0Z - проекция наджелудочкового гребня. Светлой изогнутой стрелкой показано переднее смещение выводного отдела правого желудочка, темными изогнутыми стрелками показаны направления развития приточного отдела правого желу дочка, пунктирной изогнутой кривой - граница между приточным и выводным отделами ПЖ на разных этапах его роста, малые черные стрелки - направление роста выводного отдела правого и левого желудочков. Линии 0 - Ад -А5 показывают раскрытие и изменение ориентации бульбовентрикулярного окна.
будет зад небоковая левожелуд очковая артерия в будущем зрелом сердце. Абсолютно идентично и формирование типов кровоснабжения предсердия, обусловленное разры вом задней части вокругпредсердного круга (С) (см. рис. 9,Б).
Если разрыв вокругжелудочкового круга (Е) происходит правее креста сердца (или задней межжелудочковой борозды), формируется левый тип доминантности, ле вее - правый. Если целостность этого круга сохранена за счет вокругпредсердного кру га, формируется сбалансированный тип кровоснабжения миокарда. Как свидетельст вует анализ собственного материала, несогласующегося, кстати, с большинством дан ных отечественных исследователей (Р. Н. Яшвили, 1962; А. Т. Юрченко, 1969; С. В. Са мойлова, 1970; Л.А.Арсентьева, 1972; В. В. Кованое и Т. Н.Аникина, 1974; И. X. Рабкин, 1977), в подавляющем числе наблюдений формируется правый тип доминантнос ти. Обусловлено это, на наш взгляд, двумя факторами: во-первых, разрыв венечного сплетения правее креста сердца требует разрыва трех кругов (С, Д, Е), левее - одного (D); во-вторых, рост левого желудочка больше удаляет правые края вокругбульбарных и вокругжелудочковых кругов от его левого края, чем от правого (см. рис. 9).
38