

4 КУРС (ЭМБРИОЛОГИЯ) / Несортированное по эмбриологии / 1лекции
.pdfТакое возможно при разделении зародыша до пятых суток развития, т.е. до образования трофобласта (рис.1). Все остальные близнецы имеют общий хорион – разделение произошло после образования трофобласта. Закладка амниона заканчивается к девятым суткам эмбриогенеза: следовательно, если разделение произошло между пятыми и девятыми сутками (после образования хориона, но до образования амниона), близнецы имеют общий хорион, но разные амнионы. У человека это случается примерно в 75% монозиготных близнецов. Примерно, в 5% случаев встречается один хорион и общий амнион, т.е. разделение имело место после девятых суток. Если у таких эмбрионов еще и было неполное разделение зародышевого щитка, можно ожидать варианты сросшихся «сиамских близнецов». Следует напомнить, что способность создавать целый организм из части называется регуляцией. К ней можно отнести создание аллофенных особей. Интересен факт: аллофенные организмы возможны и у человека. Это люди содержащие половые хромосомы XX и XY в одном организме. Такое может быть при объединении двух ранних зародышей разного (или одного пола), до процесса имплантации, с успешным созданием единого организма.
Тема: Детерминация, эмбриональная регуляция и индукционные процессы в раннем развитии (из К.Г. Газаряна).
1. Основные понятия.
Одна из основных задач экспериментальной эмбриологии изучение процесса детерминации клеток и многоклеточных зачатков. Детерминация – это определение пути дальнейшего развития (или судьбы) некоторого участка зародыша (его отдельной клетки или группы клеток), который условно можно назвать элементом зародыша.
Когда в ходе развития наступает детерминация и чем она обусловлена – основные вопросы раннего развития. Ни один элемент зародыша не детерминирован «изначально», всегда можно найти период начального развития, когда судьба элемента еще не определена (не детерминирована). Существование такого периода послужило основанием для введения понятий потенция элементов и регуляция путей их развития. Потенции – это максимальные возможности элементов зародыша, т. е. те направления их развития, которые могли бы осуществиться; в норме реализуется лишь одно из них, а остальные могут быть выявлены в эксперименте. Наличие периода развития, когда элементы еще не детерминированы, показывает, что изначальные их потенции, как правило, шире, чем те, которые реализуются. Это лежит в основе явлений эмбриональных регуляций, описанных немецким эмбриологом Г. Дришем (1867-1941). Эмбриональные регуляции – это восстановление нормального хода развития целого зародыша или зачатка после его искусственного или естественного нарушения; такое восстановление достигается благодаря изменению путей развития отдельных элементов. Эмбриональные регуляции не только свидетельствуют о существовании периода недетерминированности, но и показывают, каким образом совершается переход из этого состояния в состояние детерминации. Детерминация, как правило, идет от целого к частям: сначала детерминируется целый зачаток зародыша, но судьба отдельных его элементов (клеток) еще не определена, затем постепенно или путем скачкообразных переходов детерминируются отдельные элементы. Чем определяется судьба элементов – вопрос сложнее, нежели – когда она определяется. Лишь в сравнительно немногих случаях на первый вопрос удалось дать ответ, да и то он не является исчерпывающим. Речь идет об опытах, в которых было показано детерминирующее (или индуцирующее) воздействие одного эмбрионального зачатка на другой. Опыты по индуцирующим воздействиям привели к необходимости ввести еще одно понятие: компетенция, элемента зародыша. Компетенция – это способность элемента воспринимать индуцирующие воздействия. Как правило, компетенция приходится на заключительную фазу периода индетерминации («не детерминации»).
2. Эксперименты по выявлению эмбриональных регуляций. Регуляционные и мозаичные яйца.
Эксперименты по выявлению сроков детерминации в раннем развитии разных видов животных привели к понятию эмбриональных регуляций. В экспериментах использовали следующие основные методические приемы: 1) удаление части материала еще не дробящейся яйцеклетки (отсасывание ооплазмы, отрыв фрагмента яйцеклетки путем центрифугирования), отдельных бластомеров на стадиях дробления, участков бластулы или гаструлы, 2) добавление избыточного материала путем слияния яйцеклеток или зародышей, сращивания бластомеров и т. п., 3) перемешивание материала яйцеклетки или зародыша (центрифугирование ооплазмы, разъединение и произвольное соединение бластомеров). При этом выяснялось, изменяются ли пути развития элементов зародыша. Во многих случаях оказалось, что, несмотря на эти экспериментальные воздействия, путем эмбриональных регуляций возникали целые нормальные зародыши (что было бы невозможно, если хотя бы некоторые элементы не изменяли пути своего развития).
До начала дробления на яйцеклетках было сделано множество опытов, в которых от них отрывали часть содержимого путем сильного центрифугирования, отсасывали часть ооплазмы пипеткой или, наоборот, сливали две или несколько яйцеклеток в одну. За очень редкими исключениями (яйца насекомых, где не регулировалась уничтоженная или отторгнутая половая плазма) отрыв или отсасывание даже значительного фрагмента (1/4-1/3 яйцеклетки) не препятствовало дальнейшему нормальному развитию целого, хотя и уменьшенного в размерах организма. Такие результаты были получены на яйцеклетках многощетинковых червей, моллюсков, асцидий, кишечнополостных, иглокожих. Следовательно, накопленные в оогенезе вещества не предраспределены в яйцеклетке каким-то единственно возможным для развития образом. Этот вывод подкреплен также большим количеством опытов по разделению, сращиванию и полной «перетасовке» бластомеров дробящихся яиц. На основе полученных данных немецкий эмбриолог В. Ру разделил яйцеклетки всех изученных в этом отношении животных на две группы: регуляционные и мозаичные. К первой группе были отнесены яйцеклетки, способные к регуляции, ко второй – яйцеклетки мозаичные, состоящие из жестко детерминированных участков цитоплазмы или бластомеров, не способных изменять ход своего развития и тем самым осуществлять регуляцию. Это связано с тем, что, в мозаичных – РНК синтезируется еще в оогенезе и строго определяет дифференцировку бластомеров. Мозаичный тип развития характерен для животных со спиральным дроблением. При регуляционном типе развития исходных, материнских РНК не хватает для конкретного дальнейшего пути развития бластомеров. Здесь при дифференцировке имеют место сложные эмбриональные взаимоотношения структур зародыша. (Деление яиц на мозаичные и регуляционные условно: накопилось много экспериментальных
данных показывающих, что для всякого развития в некоторой мере свойственна как регуляция так и мозаичность).
Одними из наиболее «регуляционных» являются яйца многих гидромедуз, например Aegineta. У этого вида нормальная особь развивается из 1/32 яйца (из одного бластомера на стадии 32 бластомеров). Таковы и яйца морских ежей, у которых нормальный зародыш развивается из меридионального сектора в 1/2, 1/4 или даже1/8 яйца на любой стадии дробления до бластулы включительно – лишь бы этот сектор включал в равной пропорции как анимальные, так и вегетативные зоны. Из яиц с резким преобладанием анимального или вегетативного материала нормальные зародыши не получаются: из анимальных фрагментов образуются покрытые ресничками бластулы, далее не развивающиеся, а из вегетативных участков – напротив, зародыши с преобладанием энтодермального материала и недоразвитыми эктодермальными частями. Эти данные указывают на какую-то неоднородность дробящегося яйца по анимально-вегетативной оси.
Нормальных личинок морского ежа удается получить даже из разделенных, а затем перемешанных в беспорядке бластомеров. При этом ход регуляционного процесса еще более, чем в рассмотренном выше примере, отклоняется от нормального развития. Например, кишечник образуется не инвагинацией, а шизоцельным путем из плотной клеточной массы. Иногда даже личиночный скелет возникает раньше, чем покровы личинки, и последние затем образуются вокруг него. Это удивительное свойство развития происходить совершенно разными путями к одному и тому же конечному результату называется эквифинальностью. Эквифинальность можно рассматривать как одно из важнейших свойств регуляционных процессов.
При рассмотрении регуляционной способности яиц амфибий, борозда первого деления дробления у них либо рассекает серый серп, либо лежит в стороне от него. Если отделить друг от друга два первых бластомера путем перешнуровки, то в первом случае из обоих разовьются совершенно нормальные зародыши. Во втором случае целый организм образуется только из бластомера, содержащего серый серп, а из другого получится так называемый «брюшной комок»: неоформленный материал энтодермы, одетый также недифференцированной эктодермой. Таким образом, для нормального развития достаточна хотя бы половина серого серпа. Именно этот факт, открытый немецким эмбриологом Г. Шпеманом, подвел его к обнаружению индукционных явлений.
При культивировании на искусственных средах яйцеклеток млекопитающих открылись возможности испытания и даже использования на практике их регуляционных способностей. Они оказались очень высокими. Например, нормальный зародыш мыши развивается из каждого бластомера двух клеточной стадии. Кроме того, нормальное развитие наблюдается и после слияния 2-3 эмбрионов.
Яйца асцидий, гребневиков, круглых червей и спирально дробящихся форм традиционно относили к мозаичным. Действительно, у первых трех из перечисленных видов изолированные бластомеры дают те же зачатки (и не больше), что и при нормальном развитии. Очень четко это показано у гребневиков, обладающих в норме восемью рядами гребных пластинок, расположенных по меридианам через равные углы. При развитии зародыша из 1/2 яйца получается только 4 ряда гребных пластинок, при развитии из 1/4 только 2 ряда и т. д. Отсюда можно сделать вывод, что у этих форм в период ооплазматической сегрегации достигается достаточно жесткая, необратимая расстановка структур. У спирально дробящихся яиц один из бластомеров может иметь полярную лопасть. Формообразовательные потенции таких бластомеров резко повышены по сравнению с бластомерами, лишенными полярной плазмы. Потомство последних не может образовать мезодерму и ее производные. Бластомеры с полярной плазмой после своей изоляции, по крайней мере, до стадии 4 бластомеров включительно, способны образовать полноценный зародыш. Если материал полярной плазмы равномерно распределить между двумя первыми бластомерами, то из каждого получится целый зародыш, в результате чего возникают срощенные двойники. Значит, во всех остальных отношениях, кроме наличия или отсутствия полярной лопасти, бластомеры яиц спирально дробящихся форм равноценны.
В полной мере это относится к яйцам Spiralia, лишенным полярной плазмы или лопастей. Целый зародыш у них может быть получен из любой 1/2 яйца. Положение дорсальной и вентральной сторон, а, следовательно, и судьба левых и правых бластомеров изначально не определены, а зависят от положения веретена первого деления дробления. Это означает, что детерминирующую роль у спирально дробящихся форм играет не столько предшествующая дроблению ооплазматическая сегрегация, сколько сам процесс дробления.
Регуляционные способности бластул – ранних гаструл амфибий – испытывались в опытах по пересадкам сравнительно небольших участков из одного положения на зародыше в другое и их вырезанию из зародыша и помещению в физиологический раствор (эксплантация). Хотя эти опыты неравноценны, все они подтверждают способность практически всех участков зародыша изменять путь своего развития, способны развиваться в разных направлениях и при пересадках дифференцироваться в соответствии со своим новым положением. Основные выводы, следующие из опытов по регуляциям: 1) они показали до какого момента развития судьба тех или иных частей зародыша еще не детерминирована, и тем самым помогают выявить, на каком этапе осуществляются основные детерминирующие процессы. Показано, что такие процессы могут происходить как в оогенезе, так и в период дробления и еще позже; 2) явления регуляций позволили выявить целостный контроль над детерминацией элементов, т. е. механизмы, подчиняющие судьбу части целому. Г. Дриш сформулировал этот вывод
так: «Проспективное значение каждого элемента, ( т . е . то, как он будет развиваться дальше) есть функция его положения в целом».
3. Явление регуляций в нормальном развитии.
Изложенные данные показали, что в условиях экспериментов клетки зародыша способны изменить свою судьбу. Исследователей беспокоил вопрос – насколько точно детерминирована судьба клеток при нормальном развитии, или у всех ли зародышей данного вида все эмбриональные клетки занимают в точности одинаковые места. Многочисленные наблюдения неопровержимо показывают, что такая «поклеточная точность» встречается лишь в виде исключений. Она установлена у коловраток, круглых червей. Они действительно состоят из строго определенного числа клеток, каждая из которых расположена на определенном месте и выполняет определенную функцию. У подавляющего числа организмов клеточная точность утрачивается либо в ходе дробления, либо на последующих стадиях. Так, у кишечнополостных при анархическом дроблении положение каждого бластомера явно случайно. Можно сказать, что анархическое дробление – самой природой поставленный опыт по перемешиванию бластомеров. Но и у форм с детерминированным дроблением встречаются как неточности в расположении бластомеров, не препятствующие дальнейшему развитию, так и просто несколько разных, но равноправных способов их взаимного расположения (например, у круглых червей).
Приведенные факты показывают, что клетки отнюдь не движутся по жестко заданным наперед «формообразовательным траекториям»: их поведение может быть более или менее неодинаковым и даже независимым друг от друга. Однако, такая многовариантность не сбивает развитие зародыша с основного пути к конечной цели. Не только в эксперименте, где искусственно нарушается нормальный ход развития, но и в естественных условиях ход развития клеток регулируется согласно их положению в целом. Такой гибкий регуляторный характер развития имеет существенный адаптивные преимущества: если какое-либо одно звено будет подавлено или полностью выпадет, сохраняются шансы на достижение нормального конечного результата. С другой стороны, в экстремальных условиях резерв изменчивости может стать источником эволюционных преобразований.
4. Первичная эмбриональная индукция у амфибий.
В опытах по перешнуровке яиц амфибий было показано, что целый зародыш возникал лишь из тех бластомеров, которые включали в себя хотя бы часть материала серого серпа. Из бластомеров, лишенных этого материала, не развивалось ничего, кроме недифференцированной энтодермы и покровов тела, и таким образом осевые органы нацело отсутствовали. Между тем в норме из материала серого серпа образуются лишь хорда, прехордальная пластинка и часть осевой
мезодермы (хордомезодермы). Чем же объяснить отсутствие нервной системы при развитии из бластомеров, лишенных материала серого серпа? Г. Шпеман сделал смелое предположение, что хотя она и не возникает из материала серого серпа, но индуцируется им, т. е. формируется из индифферентной эктодермы под его влиянием.
Для проверки этого предположения надо было зачаток, развивающийся из материала серого серпа, т. е. хордомезодерму, привести в контакт с таким материалом, из которого нервная система в норме никогда не развивается, например, с эктодермой вентральной стороны тела. Таким образом, следовало пересадить зачаток хордомезодермы в вентральную область зародыша. При этом надо было исключить возможность случайного занесения вместе с хордомезодермой участка презумптивного материала нервной системы. Между тем это нелегко сделать, потому что на карте презумптивных зачатков материал нервной системы прилегает к материалу хордомезодермы, и никакой видимой границы между ними на живом зародыше нет.
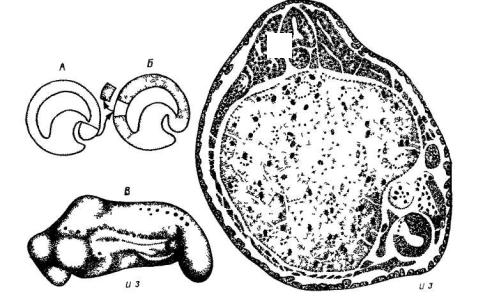
Для отведения возможных возражений Г. Шпеман применил метод гетеропластики. Он взял трансплантат зачатка хордомезодермы (дорсальной губы бластопора) от зародыша гребенчатого тритона, ткани которого были лишены пигмента, и пересадил его под брюшную эктодерму зародыша обыкновенного тритона с пигментированными тканями. После окончания опыта по распределению пигментированных и непигментированных клеток на срезах тканей зародыша можно было видеть, что произошло от трансплантата, а что возникло из тканей хозяина (рис. 34). Результаты этого замечательного опыта, поставленного в 1924 году были таковы. Примерно через сутки после пересадки дорсальной губы на брюшной стороне зародыша хозяина развились отчетливые осевые структуры: нервная трубка, сомиты, хорда, а также зачатки эмбриональных почек. Изменилась даже прилежащая энтодерма, в ней появилась полость кишечника. Анализ материала,
Г
Рис. 34. Схема опыта Г. Шпемана и Г. Мангольд (из К.Г. Газаряна). А и Б – операция по пересадке хордомезодермального зачатка; В – внешний вид полученного зародыша, Г – разрез через зародыш после операции. И.з. – индуцированный зародыш на вентральной стороне.
вошедшего в состав всех этих структур, показал, что большинство их возникло из клеток зародыша хозяина: почти вся нервная трубка, а также часть мезодермальных тканей. Таким образом, произошла настоящая индукция – глубокое изменение свойств местной ткани, которая в норме дала бы только покровную эктодерму и, возможно, некоторое количество мезенхимных клеток. Сама же пересаженная хордомезодерма, которую теперь называют индуктором, образовала, как и следовало ожидать, хорду, часть мезодермы, а также небольшой участок нервной трубки. Очень интересный побочный результат этого опыта состоит в том, что структура органов осевого комплекса оказалась чрезвычайно точной, хотя клетки трансплантата и хозяина заняли в них в общем случайные положения. Фактически в опыте Г. Шпемана произошла не только индукция, но и регуляция, аналогичная тем, что были в опытах с перемешиванием клеточного материала. Но главное внимание исследователей всего мира вызвал ясно доказанный факт индукции одной закладки другой, а именно – нервной трубки хордомезодермой. Это явление позже назвали первичной эмбриональной индукцией. Уже тогда были известны индукционные процессы в более позднем развитии, которые были названы вторичными и третичными; более ранних индукционных процессов у позвоночных тогда известно не было. Однако, как было показано позже, есть и более ранние индукционные процессы.
Надо заметить, что сам Г. Шпеман использовал термин не индукция, а организация и назвал хордомезодерму первичным организатором или организационным центром, а не индуктором, как теперь принято Эти различия в терминологии, которые могут показаться незначительными, отражают сложную и драматическую историю дальнейших работ по этой проблеме.
5. Первичная индукция в других классах хордовых. Понятие компетенции эмбриональной ткани.
Уже вскоре после открытия Шпемана была продемонстрирована индукционная способность дорсального края бластодиска рыб. Несколько позже было показано, что и гензеновский узелок у птиц (который надо считать гомологом дорсальной губы бластопора амфибий) проявляет индукционное действие: будучи пересажен в любое место зародышевого диска, он вызывает образование над собой целого дополнительного зародыша. Был проведен очень тонкий опыт по имплантации в бластоцель ранней гаструлы ланцетника дорсальной губы от другого зародыша ланцетника. Он оказал на окружающие ткани совершенно такое же индуцирующее действие и вызвал образование
дополнительной нервной трубки.
Позже было отмечено, что опыты по индукции у ланцетника ставились при температуре несколько выше нормальной для этих животных и что при нормальной температуре нервная система у них может развиваться и без воздействия индуктора. Если это верно, то можно сделать вывод – индуктор требуется ланцетнику лишь в качестве дополнительной «подстраховки» в экстремальных условиях.
Интересными оказались результаты опытов на зародышах асцидий. Основные зачатки у этих животных детерминируются еще в период дробления. Картирование яйца асцидий на стадии 8 бластомеров показало, что, как материал хордомезодермы, так и основная часть нейрального материала локализованы в одном и том же заднем вегетативном бластомере и поэтому недоступны разделению. Однако небольшая часть нейрального материала, формирующего головной ганглий, расположена в заднем, анимальном бластомере, лежащем как раз над задним вегетативным. Существуют ли индукционные влияния заднего вегетативного бластомера на задний анимальный? Что бы это проверить, анимальный ярус бластомеров поворачивали на 180° так, чтобы задний анимальный бластомер терял контакты с задним вегетативным, но взамен последний оказался бы в контакте с передним анимальным бластомером, из которого в норме возникал только покровный эпителий. По аналогии с амфибиями в переднем анимальном бластомере следовало бы ожидать возникновения индуцированного головного ганглия; однако этого не произошло. Быть может, у асцидий головной ганглии возникает совершенно автономно от хордомезодермального материала? Тогда он должен был бы развиться из перемещенного задне-анимального бластомера. Но не наблюдалось и этого: ганглий нигде не появился. Из этого интересного опыта следуют сразу два вывода: 1) для возникновения головного ганглия у асцидий требуется воздействие хордомезодермального материала, которое можно уподобить индукционному, но 2) никакой материал, кроме того (задне-анимального), который в норме образует этот ганглий, воспринять эти воздействия не в состоянии. Этот результат хорошо объясним, если вспомнить о компетенции. Изменение хода развития возможно лишь в том случае, если компетентная к образованию некоторой закладки область шире, чем область, из которой эта закладка в норме развивается. Именно так происходит у амфибий: к формированию нервной системы компетентна вся покровная эктодерма. Аналогично дело обстоит во всех других классах хордовых, кроме асцидий. У последних же область компетенции не шире размеров нормальной закладки, отчего индукцию из другого материала получить не удается. В дальнейшем в ходе эволюции хордовых произошло расширение областей и удлинение срока компетенции. И то и другое – признак существенного эволюционного прогресса.
Компетенция эмбриональных тканей, по всей видимости, служит отражением каких-то относительно автономных внутренних процессов в

клетках, характер и темп которых трудно или просто невозможно изменить экспериментальными воздействиями. Например, независимо от того, произойдет ли гаструляция или первичная индукция, компетенция к восприятию действия индуктора возникает в эмбриональной эктодерме к началу стадии гаструлы и прекращается к концу гаструляции. Иногда говорят об автономных «часах компетенции» в тканях, связывая компетенцию с какими-то внутриклеточными синтетическими процессами. Однако, сведения об этом еще недостаточны.
6. Возникновение индукционных свойств в ходе развития.
Голландский эмбриолог П. Ньюкуп вырезал из бластулы тритона область презумптивной мезодермы (зону III на рис. 35, А) и сращивал то, что осталось, – энтодерму (зону IV) с эктодермой (зоны I и II). Такие «укороченные» в анимально-вегетативном направлении зародыши развивались нормально; хордомезодерма с нормальной способностью к индукции восстанавливалась из той части эктодермы, которая оказалась теперь в контакте с энтодермой. Ньюкуп полагает, что продемонстрировал еще более ранний индукционный процесс, чем первичная индукция Шпемана, – активацию в эктодерме мезодермальных свойств под влиянием контакта с энтодермой (этот предполагаемый процесс показан стрелками на рис. 35,
Б).
Рис. 35. Схема опыта П. Ньюкупа по индукции мезодермы из презумптивной эктодермы под влиянием энтодермы у зародышей хвостатых амфибий (из К.Г. Газаряна). А – бластула до операции; I и II – презумптивная эктодерма, III – презумптивная мезодерма, IV – презумптивная энтодерма; Б – индукция в эктодерме мезодермальных свойств (стрелки) после удаления зоны III.
Таким образом, в опытах Ньюкупа свойство быть индуктором показано на материале, вовсе не происшедшем из зоны серого серпа. Шведскому эмбриологу С. Левтрупу удалось заново получить серый серп с индукционными свойствами в вентральной половине яйца амфибий, помещенного в условия довольно крутого градиента аэрации (снабжения кислородом): серп возник на более аэрированной стороне. Напомним, что