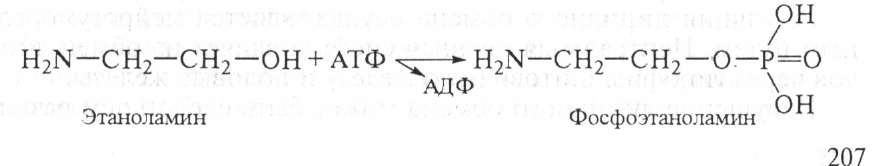2. Если синтезируется много сн3—со—КоА, а энергии для синтеза жира недостаточно, то образуется активированная ацетоуксусная кислота:
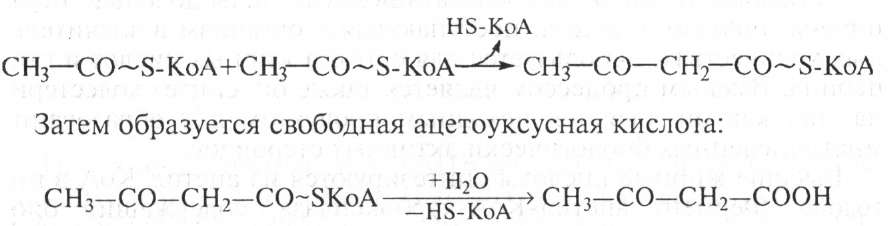
Ацетоуксусная кислота может превращаться в β-оксимасляную кислоту или ацетон, как указано выше. Кетоновые тела синтезируются в печени и поступают в кровь. В нормальных условиях β-оксимасляная, ацетоуксусная кислоты окисляются в мышцах и служат источником энергии. Однако при нарушениях обмена (сахарный диабет), когда использование углеводов затруднено и усиливается окисление липидов, при длительном недостатке углеводов происходит чрезмерный биосинтез кетоновых тел с образованием ацетона. При этом возникает кетонемия (ацидоз) и нарушается нормальное функционирование организма.
Так, при избыточном кормлении коров концентратами, при повышенном маслянокислом брожении в преджелудках и после голодания в крови жвачных резко возрастает уровень кетоновых тел, недоокисленных продуктов жирового обмена — развиваются кетозы. Если у клинически здоровых животных уровень кетоновых тел равен 0,06...0,07 г/л, то при кетозах их содержание в крови достигает 0,48...0,50 г/л, а в моче —до 2,5...3 г/л (норма 0,09... 0,1 г/л). При кетозах возникает ацидоз, уменьшается содержание
глюкозы в крови, резко падает продуктивность животных, происходит перерождение паренхиматозных органов и в тяжелых случаях наступает смерть.
Окисление ненасыщенных жирных кислот. Сначала ненасыщенные жирные кислоты превращаются в соответствующую насыщенную жирную кислоту. Затем они окисляются по β-углеродному атому.
Окисление жирных кислот с нечетным числом углеродных атомов происходит так же, как и при окислении жирных кислот с четным числом атомов углерода. Отличие состоит в том, что в конечном счете образуется пропионил-КоА, который вначале подвергается карбоксилированию, в результате образуется активированная янтарная кислота (сукцинил-КоА), которая окисляется в цикле трикарбоновых кислот.
12.3. Биосинтез липидов
Способность животных запасать полисахариды довольно ограничена, поэтому глюкоза, поступающая в организм в значительных количествах, используется для
синтеза жирных кислот и глицерина. Важным процессом является также биосинтез холестерина, так как он является исходным продуктом для образования многочисленных биологически активных стероидов.
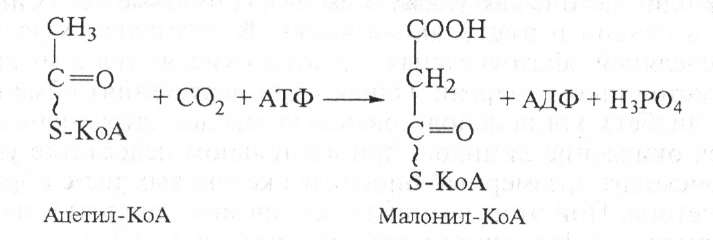
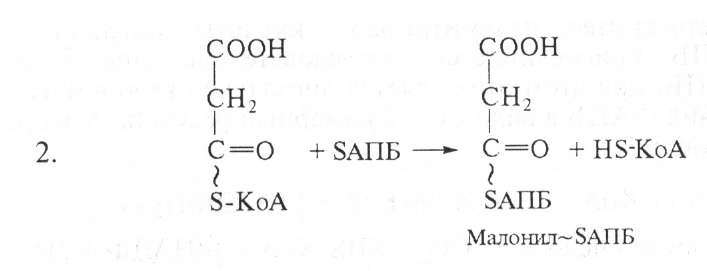

На следующем этапе ацетил-БАПБ и малонил-БАПБ взаимодействуют с образованием ацетоацетил -SАПБ:
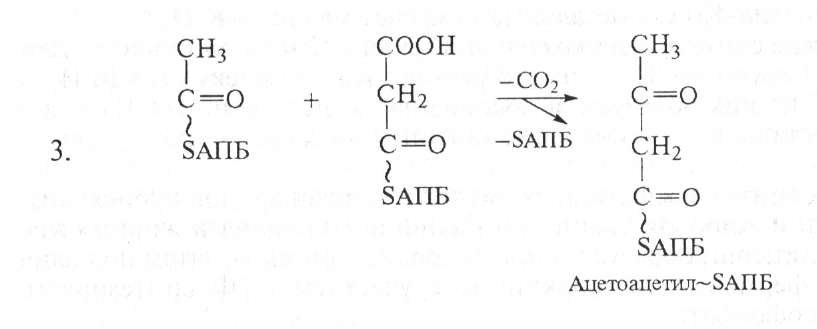
После этого ацетоацетил-SАПБ восстанавливается по β-углеродному атому с участием НАДФН2, в результате образуется β-гидроксибутирил-SАПБ. Затем к ацетилпереносящему белку (АПБ) присоединяются активированная уксусная кислота и малонил.
β-Гидроксибутирил-БАПБ под действием фермента β-гидро- кси-АПБ-дегидратазы теряет молекулу воды (5) и превращается в кротонил-SАПБ; последний с участием НАДФН2 восстанавливается в бутирил-SАПБ. Далее цикл реакции повторяется.
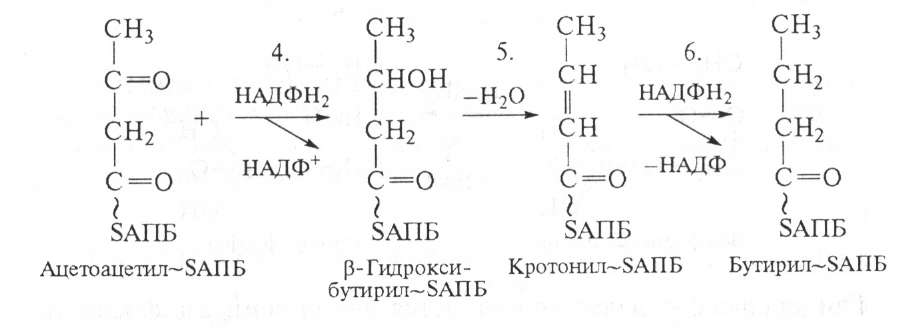
Если идет синтез пальмитиновой кислоты (С16), то к бутирил-SАПБ присоединяется последовательно еще 7 раз малонил-SАПБ, при этом отщепляется дистальная карбонильная группа малонил-SАПБ в виде СО2. Суммарный результат этих реакций следующий:
Ацетил-КоА + 7 малонил-КоА + 14НАДФН2
→пальмитиновая кислота + 7СO2 + 8hs-kOa + 14НАДФ + 7Н2O
В этом процессе семь молекул С02 образуется за счет свободных карбоксильных групп семи молекул малонил-КоА. Из 16 углеродных атомов пальмитиновой кислоты 2 атома образуется за счет ацетил-КоА, а остальные — за счет малонил-КоА.
В ходе синтеза 7 карбоксильных групп —С=0— восстанавливаются из 7 групп —СН2—, на это расходуется 14 молекул НАДФН2: за счет 7 из них образуются водородные атомы групп —СН2—, а за счет остальных 7 — кислород карбонильных групп превращается в воду.
Биосинтез фосфатидных кислот и триглицеридов происходит в печени и жировой ткани при наличии глицерина и жирных кислот. Глицерин образуется при гидролизе липидов, затем под влиянием фермента глицерокиназы с участием АТФ синтезируется глицерофосфат:
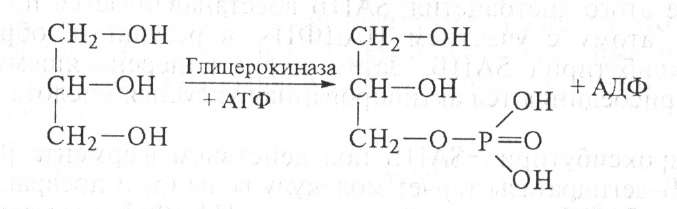
Глицерофосфат может образоваться также из диоксиацетонфосфата, возникшего при окислении глюкозы путем восстановления НАДН2:
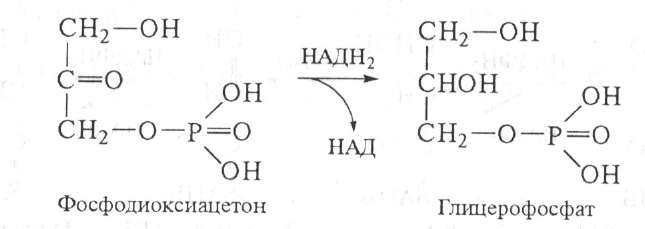
Глицерофосфат далее используется для биосинтеза фосфатидной кислоты. Реакция катализируется фосфатидсинтетазой:
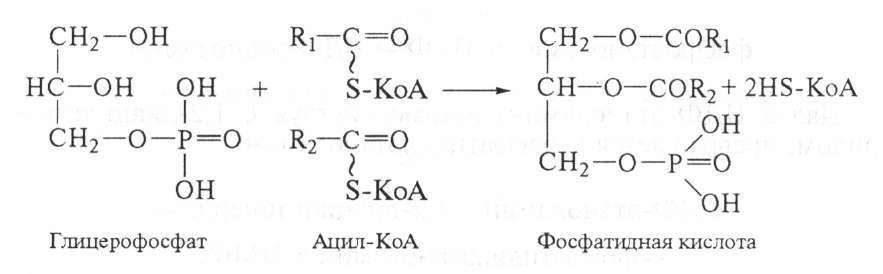
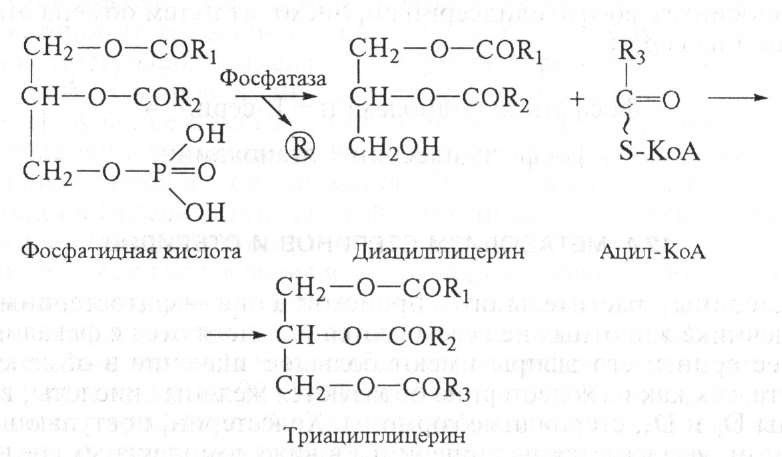
В фосфатидной кислоте в двух позициях спиртовые группы глицерина соединены сложноэфирной связью с остатками жирной кислоты (чаще всего с 16-м и 18-м углеродными атомами), а в одной позиции — с фосфорной кислотой. Триацилглицериды синтезируются из фосфатидной кислоты. Вначале от нее отделяется фосфорная кислота, при этом образуется диацилглицерин. Далее последний реагирует с третьей молекулой активированной жирной кислоты (ацил-КоА) и образуется триацилглицерин:
Биосинтез фосфоглицеридов происходит в эндоплазматических мембранах клетки. Фосфатидилэтаноламин синтезируется в следующей последовательности: вначале фосфорилируется с участием АТФ и фермента этаноламинкиназы: