

12.1. Переваривание липидов в пищеварительном тракте
В полости рта липиды подвергаются лишь механической обработке. В желудке присутствует небольшое количество липазы, которая гидролизует жиры. Малая активность липазы желудочного сока связана с кислой реакцией содержимого желудка. Кроме того, липаза может влиять только на эмульгированные жиры, а в желудке условия для образования эмульсии жира отсутствуют. Только у детей и у моногастричных животных липаза желудочного сока играет важную роль в переваривании липидов.
Кишечник является основным местом переваривания липидов. В двенадцатиперстной кишке на них воздействуют желчь печени
и сок поджелудочной железы, одновременно происходит нейтрализация кишечного содержимого (химуса). Эмульгируются жиры под действием желчных кислот. В состав желчи входят: холевая, дезоксихолевая (3,12-дигидроксихолановая), хенодезоксихолевая (3,7-дигидроксихолановая) кислоты, натриевые соли парных желчных кислот: гликохолевой, гликодезоксихолевой, таурохолевой, тауродезоксихолевой. Они состоят из двух компонентов: холевой и дезоксихолевой кислот, а также глицина и таурина:
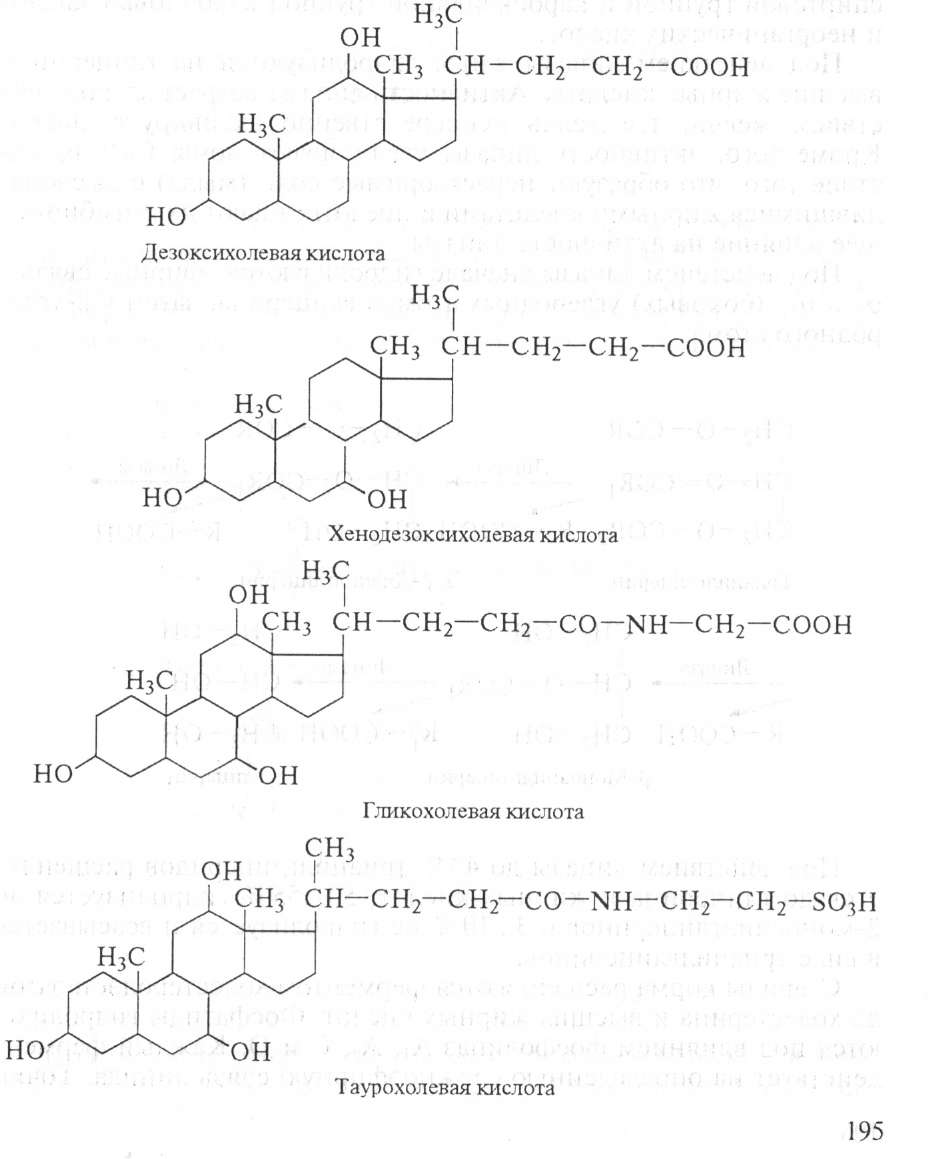
Соли желчных кислот хорошо эмульгируют жиры. При этом увеличивается площадь соприкосновения ферментов с жирами и увеличивается действие фермента. Недостаточность синтеза желчных кислот или задержка их поступления нарушает эффективность действия ферментов. Жиры, как правило, всасываются после гидролиза, но часть тонко эмульгированных жиров всасывается через стенку кишечника и переходит в лимфу без гидролиза.
Эстеразы (липазы) разрывают в жирах эфирную связь между спиртовой группой и карбоксильной группой карбоновых кислот и неорганических кислот.
Под действием липазы жиры гидролизуются на глицерин и высшие жирные кислоты. Активность липазы возрастает под действием желчи, т. е. желчь непосредственно активирует липазу. Кроме того, активность липазы увеличивают ионы Са+2 вследствие того, что образуют нерастворимые соли (мыла) с высвободившимися жирными кислотами и предотвращают их ингибирую- щее влияние на активность липазы.
Под действием липазы вначале гидролизуются эфирные связи у а- и а,- (боковых) углеродных атомов глицерина, затем у -углеродного атома:
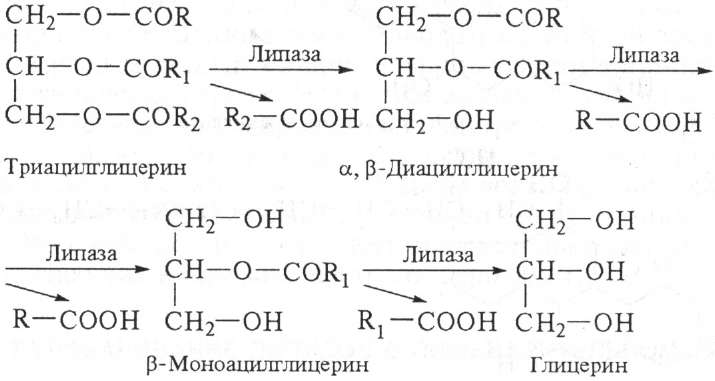
Под действием липазы до 40 % триацилглицеридов расщепляется до глицерина и жирных кислот, 50...55 % гидролизуется до 2-моноацилглицеринов и 3...10 % не гидролизуется и всасывается в виде триацилглицеринов.
Стериды корма расщепляются ферментом холестеролэстеразой до холестерина и высших жирных кислот. Фосфатиды гидролизуются под влиянием фосфолипаз Аь А2, С и D. Каждый фермент действует на определенную сложноэфирную связь липида. Точки
приложения фосфолипаз представлены на схеме:

![]()
Фосфолипазы поджелудочной железы, тканевые фосфолипазы вырабатываются в виде проферментов и активируются трипсином. Фосфолипаза А2 змеиных ядов катализирует отщепление ненасыщенной жирной кислоты в положении 2 фосфоглицеридов. При этом образуются лизолецитины с гемолитическим действием:
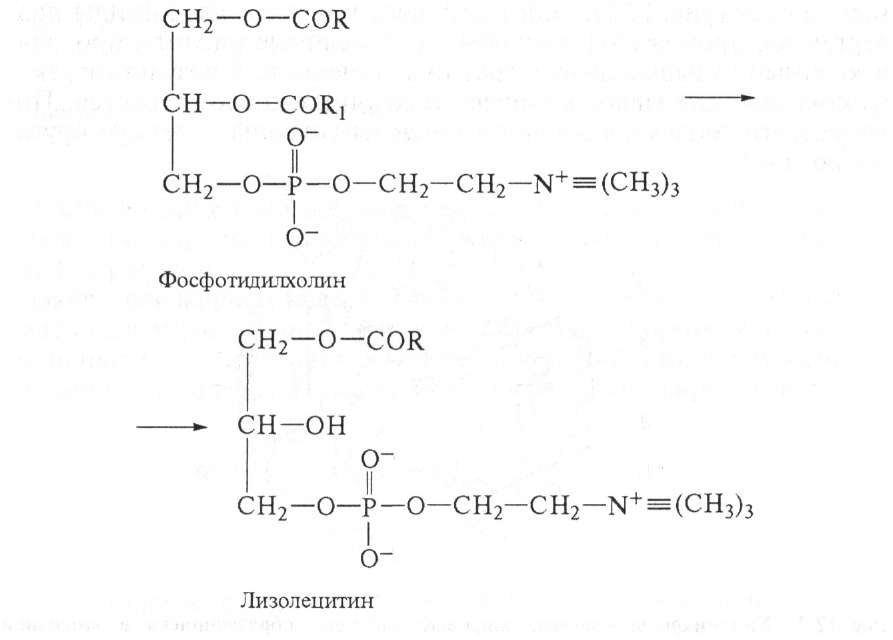
липазы А,, быстро инактивирующей лизофосфатид в результате отщепления от него остатка насыщенной жирной кислоты с превращением его в неактивный глицерофосфохолин.
Лизолецитины в малых концентрациях стимулируют дифференцировку лимфоидных клеток, активность протеинкиназы С, усиливают клеточную пролиферацию.
Коламинфосфатиды и серинфосфатиды расщепляются фосфо- липазой А до лизоколаминфосфатидов, лизосеринфосфатидов, которые далее расщепляются фосфолипазой А2. Фосфолипазы С и D гидролизуют связи холина, коламина и серина с фосфорной кислотой и остатка фосфорной кислоты с глицерином.
Всасываются липиды в тонкой кишке. Жирные кислоты с длиной цепи менее 10 углеродных атомов всасываются в неэтерифи- цированной форме. Для всасывания необходимо присутствие эмульгирующих веществ — желчных кислот и желчи.
