

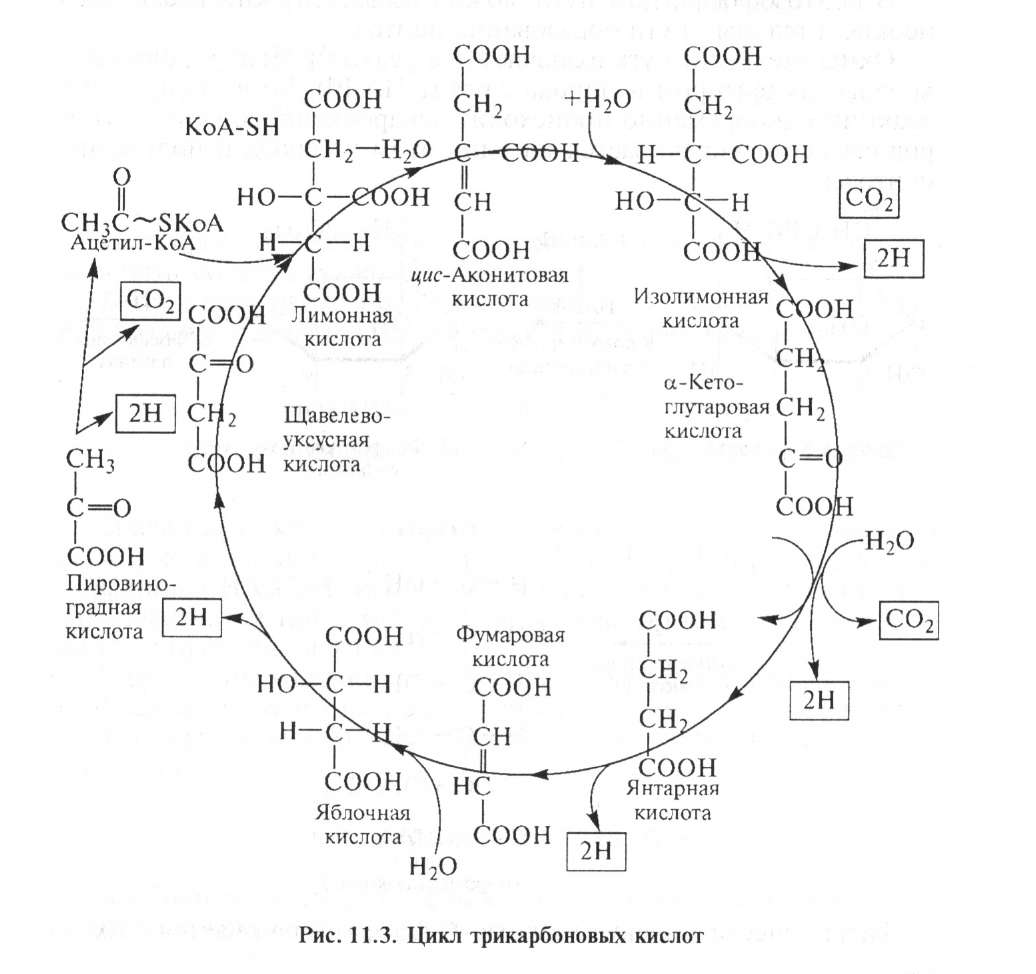
11.3. Цикл трикарбоновых кислот
Цикл трикарбоновых кислот, или цикл Кребса, — это завершающий этап распада энергетических материалов в клетке. При этом активированная уксусная кислота (ацетил-SKoA) — промежуточный метаболит углеводов, белков и липидов — окисляется до конечных продуктов — диоксида углерода, воды и энергии. Реакции цикла трикарбоновых кислот происходят во внутренних отсеках митохондрий.
Митохондрии обычно имеют форму цилиндра с закругленными концами, длиной 1...4 мкм и шириной 0,3...0,7 мкм (рис. 11.2). Число митохондрий в разных клетках различно, например в клетках печени их может быть до 2000.
Система ферментов тканевого окисления (дыхания) расположена на внутренней мембране, где происходит окисление СН3— СО~КоА до конечных продуктов — С02, Н20, с выделением энергии. В цикле трикарбоновых кислот происходит дегидрирование ди- и трикарбоновых кислот, тканевое дыхание и окислительное фосфорилирование, в этих реакциях участвуют ферменты дыхательной цепи и окислительного фосфорилирования.
Реакции цикла трикарбоновых кислот происходят в несколько этапов.


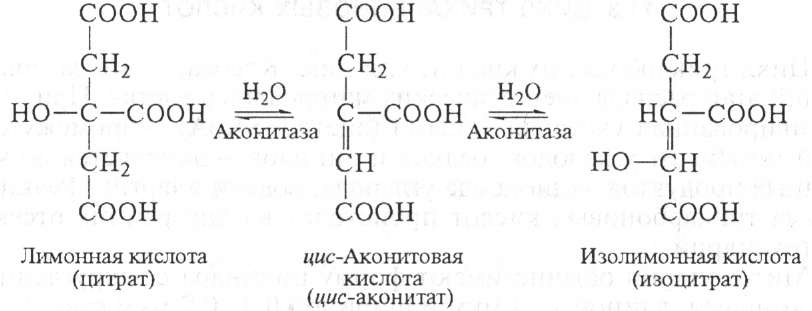
Фермент аконитаза осуществляет отнятие молекулы воды от лимонной кислоты, присоединение молекулы воды к цис-аконитовой кислоте и изомерное превращение цитрата в изоцитрат.
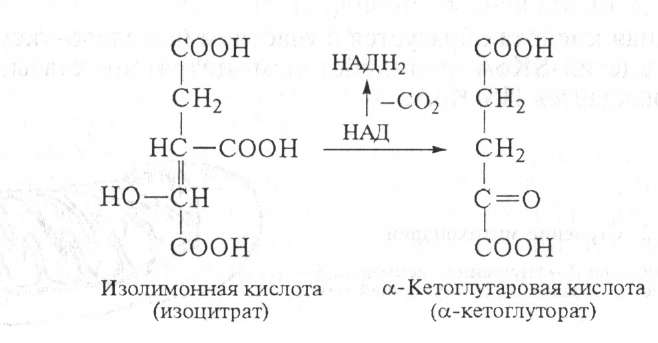
При отщеплении двух атомов водорода от изолимонной кислоты происходит восстановление НАД в НАДН2 (или в НАДФН2), который при окислении в дыхательной цепи обеспечивает синтез трех молекул АТФ.
4. а-Кетоглутаровая кислота под влиянием фермента а-кето- глутаратдегидрогеназы декарбоксилируется, в результате образуется активированная янтарная кислота — сукцинил-КоА, содержащая макроэргическую связь:
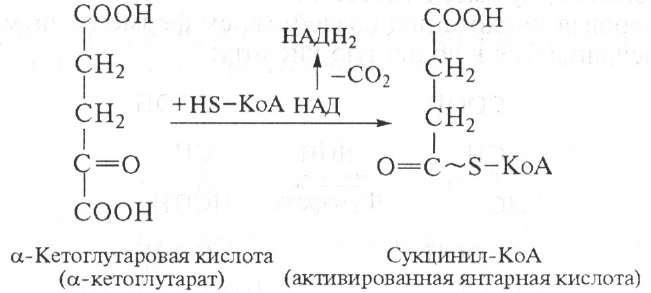
При этом НАД восстанавливается в НАДН2, который в дыхательной цепи обеспечивает синтез трех молекул АТФ.
Расщепление тиоэфирной связи сукцинил-КоА сопряжено с фосфорилированием гуанозиндифосфата (ГДФ):
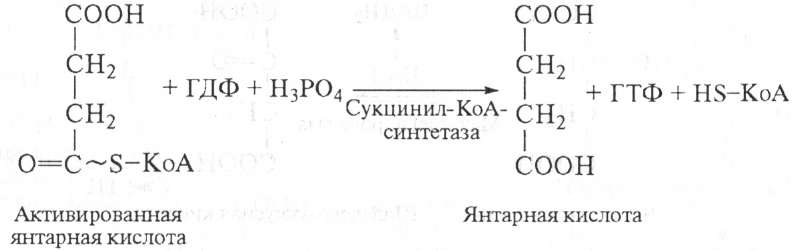
Образовавшийся при этом ГТФ вступает в реакцию перефосфорилирования с АДФ по следующей схеме:
![]()
5. Янтарная кислота под воздействием фермента сукцинатдегидрогеназы окисляется до фумаровой кислоты. Коферментом при этом служит ФАД:

ФАД • Н2 окисляется в дыхательной цепи митохондрий и обеспечивает синтез двух молекул АТФ.
6. Фумаровая кислота под воздействием фермента фумаратгидратазы превращается в яблочную кислоту:
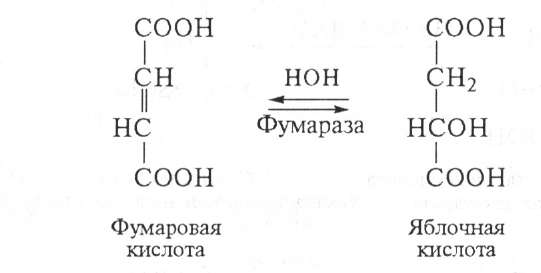
7. Яблочная кислота дегидрируется иод действием фермента малатдегидрогеназы, в результате образуется щавелево-уксусная кислота (ЩУК) и происходит восстановление НАД в НАДН2, последний в дыхательной цепи окисляется и обеспечивает образование трех молекул АТФ:
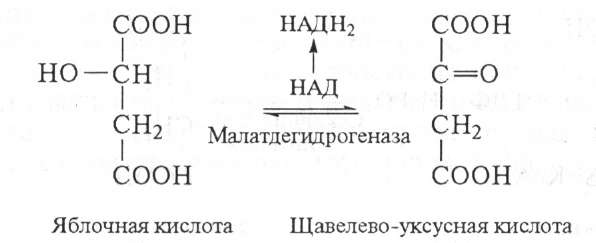
С момента образования щавелево-уксусной кислоты цикл три- карбоновых кислот замыкается и может повторяться в том же порядке: при наличии в системе ацетил-КоА ЩУК снова включается в цикл трикарбоновых кислот.
Суммарная реакция окисления ацетил-КоА в цикле трикарбоновых кислот выглядит следующим образом:
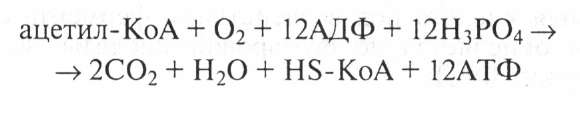
Таким образом, в одном цикле трикарбоновых кислот до конечных продуктов — СО, и Н20 — окисляется одна молекула активированной уксусной кислоты (ацетил-КоА), а энергия, высвобожденная при этом, аккумулируется в 12 молекулах АТФ.
![]()
Из них 24 молекулы АТФ синтезируются в цикле трикарбоновых кислот, 6 — при декарбоксилировании пировиноградной кислоты в ацетил-КоА, 2 — при анаэробном гликолизе, 6 — при окислении НАДН2, образующегося при субстратном окислении фосфоглицеринового альдегида в цитозоле.