

2.2 Причина сукцессий (частных).
Сукцессионные смены обычно связывают с тем, что существующая экосистема (сообщество) создает неблагоприятные условия для наполняющих ее организмов (почвоутомление, неполный круговорот веществ, самоотравление продуктами выделения или разложения и т.п.). Такие явления реальны, но не объясняют всех случаев смен экосистем. Например, в северных лесах внедрение под полог лиственных древесных сообществ ели связано, прежде всего, с тем, что последняя использует биологические свойства первых по слабому притенению почвы. Сами же почвенные условия остаются не только благоприятными для лиственных древостоев, но и постепенно улучшаются для них (идет накопление питательных веществ, уменьшается кислотность и т.п.). Не подтверждается точка зрения, что появление ели под пологом лиственных лесов и древостоев связано с тем, что в молодом возрасте этот вид требует затенения. Известно, что ель и в молодом возрасте прекрасно растет при полном освещении (значительно лучше, чем под пологом других древесных видов). Об этом, в частности, свидетельствуют многочисленные примеры создания культурных фитоценозов ели (посадкой молодых растений или посевом семян) на открытых площадях.
2.3 Человеческий фактор.
Наряду с природными факторами, причинами динамики экосистем все чаще выступает человек. К настоящему времени им разрушено большинство коренных (климаксных) экосистем. Например, степи почти полностью распаханы (сохранились только на заповедных участках). Преобладающие площади лесов представлены переходными (временными) экосистемами из лиственных древесных пород (береза, осина, реже ива, ольха и другие). Эти леса обычно называют производными, или вторичными. Они, как отмечалось выше, являются промежуточными стадиями сукцессий. К сменам экосистем ведут также такие виды деятельности человека, как осушение болот, чрезмерные нагрузки на леса. Например, в результате отдыха населения (рекреации), химических загрязнений среды, усиленного выпаса скота, пожаров и т.п. Антропогенные воздействия часто ведут к упрощению экосистем. Такие явления обычно называют дегрессиями. Различают, например, пастбищные, рекреационные и другие дегрессии. Смены такого типа обычно завершаются не климаксными экосистемами, для которых характерно усложнение структуры, а стадиями катоценоза, которые нередко заканчиваются полным распадом экосистем. Климаксные экосистемы обычно чувствительны к различным вмешательствам в их жизнь. К подобным воздействиям, кроме хвойных лесов, чувствительны и другие коренные сообщества, например, дубовые леса. Это одна из причин катастрофической гибели дубрав в современный период и замена их, как и хвойных лесов, менее ценными, но более устойчивыми временными экосистемами из березы, осины, кустарников или трав. Последнее особенно типично при разрушении степных и лесостепных дубрав.
2.3 Виды сукцессий.
Сукцессии, с которыми мы познакомились на примере лесной зоны, называют первичными по той причине, что они начинаются с исходно безжизненного пространства (субстрата). Кроме отвалов горных пород, такие сукцессии могут начинаться на песчаных обнажениях, продуктах извержения вулканов (застывшая лава, отложения пепла) и т.п. Наряду с первичными выделяют вторичные сукцессии. Последние отличаются от первичных тем, что они начинаются обычно не с нулевых значений, а возникают на месте нарушенных или разрушенных экосистем. Например, после вырубок лесов, лесных пожаров, при разрастании площадей, находившихся под сельскохозяйственными угодьями. Основное отличие этих сукцессий заключается в том, что они протекают несравненно быстрее первичных, так как начинаются с промежуточной стадии (трав, кустарников или древесных растений-пионеров) и на фоне более богатых почв. Конечно, вторичная сукцессия возможна только в тех случаях, если человек не будет оказывать сильное и постоянное влияние на развивающиеся экосистемы. В последнем случае процесс пойдет по схеме дегрессий и завершится стадией катоценоза и опустынивания территорий. Различают также автотрофные и гетеротрофные сукцессии. Рассмотренные примеры сукцессий относятся к автотрофным, поскольку все они протекают в экосистемах, где центральным звеном является растительный покров. С его развитием связаны смены гетеротрофных компонентов. Такие сукцессии потенциально бессмертны, поскольку все время пополняются энергией и веществом, образующимися или фиксирующимися в организмах в процессе фотосинтеза либо хемосинтеза. Завершаются они, как отмечалось, климаксной стадией развития экосистем. К гетеротрофным относятся те сукцессии, которые протекают в субстратах, где отсутствуют живые растения (продуценты), а участвуют только животные (гетеротрофы) или мертвые растения. Этот вид сукцессий имеет место только до тех пор, пока присутствует запас готового органического вещества, в котором сменяются различные виды организмов-разрушителей. По мере разрушения органического вещества и высвобождения из него энергии сукцессионный ряд заканчивается, система распадается. Таким образом, эта сукцессия по природе своей деструктивна. Примерами гетеротрофных являются сукцессии, имеющие место, например, при разложении мертвого дерева или трупа животного. Так, при разложении мертвого дерева можно выделить несколько стадий смен гетеротрофов. Первыми на мертвом, чаще ослабленном дереве, поселяются насекомые-короеды. Далее их сменяют насекомые, питающиеся древесиной (ксилофаги). К ним относятся личинки усачей, златок и других. Одновременно идут смены грибного населения. Они имеют примерно следующую последовательность: грибы-пионеры (обычно окрашивают древесину в разные цвета), грибы-деструкторы, способствующие появлению мягкой гнили, и грибы -гумификаторы, превращающие часть гнилой древесины в гумус. На всех стадиях сукцессий присутствуют также бактерии. В конечном счете, органическое вещество в основной массе разлагается до конечных продуктов: минеральных веществ и углекислого газа. Гетеротрофные сукцессии широко осуществляются при разложении детрита (в лесах он представлен лесной подстилкой). Они протекают также в экскрементах животных, в загрязненных водах, в частности, интенсивно идут при биологической очистке вод с использованием активного ила, насыщенного большим количеством организмов.
2.4 Общие закономерности сукцессионного процесса. Для любой сукцессии, особенно первичной, характерны следующие общие закономерности протекания процесса: 1. На начальных стадиях видовое разнообразие незначительно, продуктивность и биомасса малы, но по мере развития сукцессии эти показатели возрастают. 2. С развитием сукцессионного ряда увеличиваются взаимосвязи между организмами. Особенно возрастает количество и роль симбиотических отношений. Полнее осваивается среда обитания, усложняются цепи и сети питания. 3. Уменьшается количество свободных экологических ниш, и в климаксном сообществе они либо отсутствуют, либо находятся в минимуме. В связи с этим по мере развития сукцессий уменьшается вероятность вспышек численности отдельных видов. 4. Интенсифицируются процессы круговорота веществ, поток энергии и дыхание экосистем. 5. Скорость сукцессионного процесса в большей мере зависит от продолжительности жизни организмов, играющих основную роль в сложении и функционировании экосистем. В этом отношении наиболее продолжительные сукцессии в лесных экосистемах. Короче они в экосистемах, где автотрофное звено представлено травянистыми растениями, и еще быстрее протекают в водных экосистемах. 6. Неизменяемость завершающих (климаксных) стадий сукцессий относительна. Динамические процессы при этом не приостанавливаются, а лишь замедляются. Продолжаются динамические процессы, обуславливаемые изменениями среды обитания, сменой поколений организмов и другими явлениями. Относительно большой удельный вес занимают динамические процессы циклического (флуктуационного) плана. 7. В зрелой стадии климаксного сообщества биомасса обычно достигает максимальных, или близких к максимальным, значений. Неоднозначна продуктивность отдельных сообществ на стадии климакса. Обычно считается, что по мере развития сукцессионного процесса продуктивность увеличивается и достигает максимума на промежуточных стадиях, а затем в климаксном сообществе резко уменьшается. Последнее связывают, во-первых, с тем, что в это время максимум первичной продукции потребляется консументами, а, во-вторых, экосистема развивает чрезвычайно большую массу ассимиляционного аппарата, что ведет к дефициту освещенности, следствием чего является снижение интенсивности фотосинтеза при одновременном возрастании потерь продуктов ассимиляции на дыхание самих автотрофов. Эти положения нельзя распространять на все климаксные сообщества. Например, нет реальных предпосылок для увеличения численности гетеротрофов в хвойных лесах по сравнению с лиственными. Скорее, в последних больше потребителей зеленой продукции и, вероятнее, вспышки численности отдельных видов-фитофагов, например, насекомых. Нет также ни теоретических предпосылок, ни фактических данных, которые бы свидетельствовали, что в зрелой климаксной системе, например в еловых лесах, масса хвои достигает чрезмерно высоких значений. Это противоречит принципам адаптации к увеличению биогенной геохимической энергии организмами как условию их выживания (второй биогеохимический принцип В.И. Вернадского). Весь опыт лесоводства также свидетельствует о наиболее высокой продуктивности климаксных лесных сообществ (применительно к лесной зоне хвойных или смешанных хвойно-лиственных лесов). В противном случае, с точки зрения получения продукции (древесины), неизбежен вывод о нецелесообразности ориентации на выращивание и сохранение климаксных стадий лесов. Применительно к другим экосистемам, например луговым, можно согласиться с тем, что возможности получения продукции на климаксной стадии уменьшаются, однако не потому, что сокращается ее нарастание (прирост, продуктивность), а по той причине, что более значительная часть ее отчуждается гетеротрофами в результате образования устойчивых цепей выедания. Другими словами, продуктивность экосистем на климаксных стадиях сукцессий высока. Как правило, максимальна, вследствие более полного освоения пространства. Однако возможности снятия человеком первичной продукции лимитируются (иногда до нулевых значений) вследствие включения ее в цепи питания.
Общая структура экосистем.
Экосистемы состоят из живого и неживого компонентов, называемых соответственно биотическим и абиотическим. Совокупность живых организмов биотического компонента называется сообществом. Исследование экосистем включает, в частности, выяснение и описание тесных взаимосвязей, существующих между сообществом и абиотическим компонентом.
Биотический компонент полезно подразделить на автотрофные и гетеротрофные организмы. Таким образом, все живые организмы попадут в одну из двух групп. Автотрофы синтезируют необходимые им органические вещества из простых неорганических и делают, за исключением хемотрофных бактерий, с помощью фотосинтеза, используя свет как источник энергии. Гетеротрофы нуждаются в источнике органического вещества и (за исключением некоторых бактерий) используют химическую энергию, содержащуюся в потребляемой пище. Гетеротрофы в своем существовании зависят от автотрофов, и понимание этой зависимости необходимо для понимания экосистем.
Неживой, или абиотический, компонент экосистемы в основном включает:
1) почву или воду
2) климат.
Почва и вода содержат смесь неорганических и органических веществ. Свойства почвы зависят от материнской породы, на которой она лежит, и из которой частично образуется. В понятие климата входят такие параметры, как освещенность, температура и влажность, в большой степени определяющие видовой состав организмов, успешно развивающийся в данной экосистеме. Для водных экосистем очень существенна также степень солености.
Биотический компонент экосистем
Организмы в экосистеме связаны общностью энергии и питательных веществ. Всю экосистему можно уподобить единому механизму, потребляющему энергию и питательные вещества для совершения работы. Питательные вещества первоначально происходят из абиотического компонента системы, в который, в конце концов, и возвращаются либо в качестве отходов жизнедеятельности, либо после гибели и разрушения организмов. Таким образом, в экосистеме происходит круговорот питательных веществ, в котором участвуют и живой и неживой компоненты. Такие круговороты называются биогеохимическими циклами.
Движущей силой этих круговоротов служит, в конечном счете, энергия Солнца. Фотосинтезирующие организмы непосредственно используют энергию солнечного света и затем передают ее другим представителям биотического компонента. В итоге создается поток энергии и питательных веществ через экосистему. Необходимо еще отметить, что климатические факторы абиотического, компонента, такие, как температура, движение атмосферы, испарение и осадки, тоже регулируются поступлением солнечной энергии.
Энергия может существовать в виде различных взаимопревращаемых форм, таких, как механическая, химическая, тепловая и электрическая энергия. Переход одной формы в другую называется преобразованием энергии.
Таким образом, все живые организмы – это преобразователи энергии, и каждый раз, когда происходит превращение энергии, часть ее теряется в виде тепла. В конце концов, вся энергия, поступающая в биотический компонент экосистемы, рассеивается в виде тепла. Изучение потока энергии через экосистемы называется энергетикой экосистемы.
Фактически живые организмы не используют тепло, как источник энергии для совершения работы – они используют свет и химическую энергию.
Изучение потока энергии через экосистемы называется энергетикой экосистем.
Солнце как источник энергии
Первоисточником энергии для экосистем служит Солнце. Солнце – это звезда, излучающая в космос огромное количество энергии. Энергия распространяется в космическом пространстве в виде электромагнитных волн, и небольшая часть ее, примерно 10,5 * 106 кДж/м2 в год, захватывается Землей. Около 40 % этого количества сразу отражается от облаков, атмосферной пыли и поверхности Земли без какого бы то ни было теплового эффекта. Еще 15 % поглощаются атмосферой (в частности, озоновым слоем в ее верхних частях) и превращаются в тепловую энергию или расходуются на испарение воды. Оставшиеся 45 % поглощаются растениями и земной поверхностью. В среднем это составляет 5 * 106 кДж/м2 в год, хотя реальное количество энергии для данной местности зависит от географической широты. Большая часть энергии повторно излучается земной поверхностью и нагревает атмосферу, приблизительно две трети энергии поступает в атмосферу этим путем. И только небольшая часть пришедшей от Солнца энергии усваивается биотическим компонентом экосистемы.
Пищевые цепи и трофические уровни
Внутри экосистемы содержащие энергию органические вещества создаются автотрофными организмами и служат пищей (источником вещества и энергии) для гетеротрофов. Типичный пример - животное поедает растения. Это животное в свою очередь может быть съедено другим животным, и таким путем может происходить перенос энергии через ряд организмов – каждый последующий питается предыдущим, поставляющим ему сырье и энергию. Такая последовательность называется пищевой цепью, а каждое ее звено – трофическим уровнем. Первый трофический уровень занимают автотрофы, или так называемые первичные продуценты. Организмы второго трофического уровня называются первичными консументами, третьего – вторичными консументами и т. д. Обычно бывает четыре или пять трофических уровней и редко больше шести.
Первичные продуценты
Первичными продуцентами являются автотрофные организмы, в основном зеленые растения. Некоторые прокариоты, а именно сине-зеленые водоросли и немногочисленные виды бактерий, тоже фотосинтезируют, но их вклад относительно невелик. Фотосинтетики превращают солнечную энергию (энергию света) в химическую энергию, заключенную в органических молекулах, из которых построены ткани. Небольшой вклад в продукцию органического вещества вносят и хемосинтезирующие бактерии, извлекающие энергию из неорганических соединений.
В водных экосистемах главными продуцентами являются водоросли – часто мелкие одноклеточные организмы, составляющие фитопланктон поверхностных слоев океанов и озер. На суше большую часть первичной продукции поставляют более высокоорганизованные формы, относящиеся к голосеменным и покрытосеменным. Они формируют леса и луга.
Первичные консументы
Первичные консументы питаются первичными продуцентами, т. е. это травоядные животные. На суше типичными травоядными являются многие насекомые, рептилии, птицы и млекопитающие. Наиболее важные группы травоядных млекопитающих – это грызуны и копытные. К последним относятся пастбищные животные, такие, как лошади, овцы, крупный рогатый скот, приспособленные к бегу на кончиках пальцев.
В водных экосистемах (пресноводных и морских) травоядные формы представлены обычно моллюсками и мелкими ракообразными. Большинство этих организмов – ветвистоусые и веслоногие раки, личинки крабов, усоногие раки и двустворчатые моллюски (например, мидии и устрицы) – питаются, отфильтровывая мельчайших первичных продуцентов из воды. Вместе с простейшими многие из них составляют основную часть зоопланктона, питающегося фитопланктоном. Жизнь в океанах и озерах практически полностью зависит от планктона, так как с него начинаются почти все пищевые цепи.
К первичным консументам относятся также паразиты растений (грибы, растения и животные).
Консументы второго и третьего порядка
Вторичные консументы питаются травоядными; таким образом, это уже плотоядные животные, так же как и третичные консументы, поедающие консументов второго порядка. Консументы второго и третьего порядка могут быть хищниками и охотиться, схватывать и убивать свою жертву, могут питаться падалью или быть паразитами. В последнем случае они по величине меньше своих хозяев. Пищевые цепи паразитов необычны по ряду параметров. В типичных пищевых цепях хищников плотоядные животные оказываются крупнее на каждом следующем трофическом уровне:
Растительный материал (например, нектар) → муха → паук → землеройка → сова
Сок розового куста → тля → божья коровка → паук → насекомоядная птица → хищная птица
В типичных пищевых цепях, включающих паразитов, последние становятся меньше по размерам на каждом следующем уровне.
Редуценты и детритофаги (детритные пищевые цепи)
Существуют два главных типа пищевых цепей – пастбищные и детритные. Выше были приведены примеры пастбищных цепей, в которых первый трофический уровень занимают зеленые растения, второй – пастбищные животные и третий – хищники. Тела погибших растений и животных еще содержат энергию и «строительный материал», так же как и прижизненные выделения, например, моча и фекалии. Эти органические материалы разлагаются микроорганизмами, а именно грибами и бактериями, живущими как сапрофиты на органических остатках. Такие организмы называются редуцентами. Они выделяют пищеварительные ферменты на мертвые тела или отходы жизнедеятельности и поглощают продукты их переваривания. Скорость разложения может быть различной. Органические вещества мочи, фекалий и трупов животных потребляются за несколько недель, тогда как упавшие деревья и ветви могут разлагаться многие годы. Очень существенную роль в разложении древесины (и других растительных остатков) играют грибы, которые выделяют фермент целлюлозу, размягчающий древесину, и это дает возможность мелким животным проникать внутрь и поглощать размягченный материал.
Кусочки частично разложившегося материала называют детритом, и многие мелкие животные (детритофаги) питаются им, ускоряя процесс разложения. Поскольку в этом процессе участвуют как истинные редуценты (грибы и бактерии), так и детритофаги (животные), и тех и других иногда называют редуцентами, хотя в действительности этот термин относится только к сапрофитным организмам.
Детритофагами могут в свою очередь питаться более крупные организмы, и тогда создается пищевая цепь другого типа – цепь, начинающаяся с детрита:
Детрит → детритофаг → хищник
К детритофагам лесных и прибрежных сообществ относятся дождевой червь, мокрица, личинка падальной мухи (лес), полихета, багрянка, голотурия (прибрежная зона).
Приведем две типичные детритные пищевые цепи наших лесов:
Листовая подстилка → Дождевой червь → Черный дрозд → Ястреб-перепелятник
Мертвое животное → Личинки падальных мух → Травяная лягушка → Обыкновенный уж
Некоторые типичные детритофаги - это дождевые черви, мокрицы, двупарноногие и более мелкие (<0,5 мм) животные, такие, как клещи, ногохвостки, нематоды и черви-энхитреиды.
Пищевые сети
В схемах пищевых цепей каждый организм бывает представлен как питающийся другими организмами какого-то одного типа. Однако реальные пищевые связи в экосистеме намного сложнее, т. к. животное может питаться организмами разных типов из одной и той же пищевой цепи или даже из разных пищевых цепей. Это в особенности относится к хищникам верхних трофических уровней. Некоторые животные питаются как другими животными, так и растениями; их называют всеядными (таков, в частности, и человек). В действительности пищевые цепи переплетаются таким образом, что образуется пищевая (трофическая) сеть. В схеме пищевой сети могут быть показаны только некоторые из многих возможных связей, и она обычно включает лишь одного или двух хищников каждого из верхних трофических уровней. Такие схемы иллюстрируют пищевые связи между организмами в экосистеме и служат основой для количественного изучения экологических пирамид и продуктивности экосистем.
Экологические пирамиды.
Пирамиды численности.
Для изучения взаимоотношений между организмами в экосистеме и для графического представления этих взаимоотношений удобнее использовать не схемы пищевых сетей, а экологические пирамиды. При этом сначала подсчитывают число различных организмов на данной территории, сгруппировав их по трофическим уровням. После таких подсчетов становится очевидным, что численность животных прогрессивно уменьшается при переходе от второго трофического уровня к последующим. Численность растений первого трофического уровня тоже нередко превосходит численность животных, составляющих второй уровень. Это можно отобразить в виде пирамиды численности.
Для удобства количество организмов на данном трофическом уровне может быть представлено в виде прямоугольника, длина (или площадь) которого пропорциональна числу организмов, обитающих на данной площади (или в данном объеме, если это водная экосистема). На рисунке показана пирамида численности, отображающая реальную ситуацию в природе. Хищники, расположенные на высшем трофическом уровне, называются конечными хищниками.
Ч

 етвертый
трофический уровень
Третичные консументы
етвертый
трофический уровень
Третичные консументы
Т
 ретий
трофический уровень
Вторичные консументы
ретий
трофический уровень
Вторичные консументы
В
 торой
трофический уровень
Первичные консументы
торой
трофический уровень
Первичные консументы
П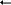 ервый
трофический
Первичные продуценты
ервый
трофический
Первичные продуценты
уровень
Пирамиды биомассы.
Неудобств, связанных с использованием пирамид численности, можно избежать путем построения пирамид биомассы, в которых учитывается суммарная масса организмов (биомассы) каждого трофического уровня. Определение биомассы включает не только учет численности, но и взвешивание отдельных особей, так что это более трудоемкий процесс, требующий больше времени и специального оборудования. Таким образом, прямоугольники в пирамидах биомассы отображают массу организмов каждого трофического уровня, отнесенную к единице площади или объема.
При отборе образцов - иными словами, в данный момент времени - всегда определяется так называемая биомасса на корню, или урожай на корню. Важно понимать, что эта величина не содержит никакой информации о скорости образования биомассы (продуктивности) или ее потребления; иначе могут возникнуть ошибки по двум причинам:
Если скорость потребления биомассы (потеря вследствие поедания) примерно соответствует скорости ее образования, то урожай на корню не обязательно свидетельствует о продуктивности, т.е. о количестве энергии и вещества, переходящих с одного трофического уровня на другой за данный период времени, например за год. Например, на плодородном, интенсивно используемом пастбище урожай трав на корню может быть ниже, а продуктивность выше, чем на менее плодородном, но мало используемом для выпаса.
Продуцентом небольших размеров, таким, как водоросли, свойственна высокая скорость возобновления, т.е. высокая скорость роста и размножения, уравновешенная интенсивным потреблением их в пищу другими организмами и естественной гибелью. Таким образом, хотя биомасса на корню может быть малой по сравнению с крупными продуцентами (например, деревьями), продуктивность может быть не меньшей, так как деревья накапливают биомассу в течение длительного времени. Иными словами, фитопланктон с такой же продуктивностью, как у дерева, будет иметь намного меньшую биомассу, хотя он мог бы поддержать жизнь такой же массы животных. Вообще популяции крупных и долговечных растений и животных обладают меньшей скоростью обновления по сравнению с мелкими и короткоживущими и аккумулируют вещество и энергию в течение более длительного времени. Зоопланктон обладает большей биомассой, чем фитопланктон, которым он питается. Это характерно для планктонных сообществ озер и морей в определенное время года; биомасса фитопланктона превышает биомассу зоопланктона во время весеннего «цветения», но в другие периоды возможно обратное соотношение. Подобных кажущихся аномалий можно избежать, применяя пирамиды энергии.
Абиотический компонент экосистемы
Абиотический, т.е. неживой, компонент экосистемы подразделяется на эдафические (почвенные), климатические, топографические и другие физические факторы, в том числе воздействие волн, морских течений и огня.
Эдафические факторы.
Наука о почвах называется почвоведением. Уже в ранних работах подчеркивалось значение почвы как источника питательных веществ для растений. Хотя мы включили почву в раздел об абиотических факторах, правильнее считать ее важнейшим связующим звеном между биотическими и абиотическими компонентами наземных экосистем. Почвой называют слой вещества, лежащий поверх горных пород земной коры. В состав почвы входят четыре важных структурных компонента: минеральная основа (обычно 50-60% общего состава почвы), органическое вещество (до 10%), воздух (15-20%) и вода (25-35%).
Минеральный скелет почвы – это неорганический компонент, который образовался из материнской породы в результате ее выветривания. Минеральные фрагменты, образующие вещество почвенного скелета различны – от валунов и камней до песчаных крупинок и мельчайших частиц глины. Скелетный материал обычно произвольно разделяют на мелкий грунт (частицы менее 2 мм) и более крупные фрагменты. Частицы меньше 1 мкм в диаметре называют коллоидными. Механические и химические свойства почвы в основном определяются теми веществами, которые относятся к мелкому грунту.
Органическое вещество почвы образуется при разложении мертвых организмов, их частей (например, опавших листьев), экскретов и фекалий. Мертвый органический материал используется в пищу совместно детритофагами, которые его поедают и таким образом способствуют его разрушению, и редуцентами (грибами и бактериями), завершающими процесс разложения. Не полностью разложившиеся органические остатки называются подстилкой, а конечный продукт разложения – аморфное вещество, в котором уже невозможно распознать первоначальный материал, - получило название гумуса. Цвет гумуса варьирует от темно-бурого до черного. В химическом плане это очень сложная смесь изменчивого состава, образованная органическими молекулами различных типов; в основном гумус состоит из фенольных соединений, карбоновых кислот и сложных эфиров жирных кислот. Гумус, подобно глине, находится в коллоидном состоянии; отдельные частицы его прочно прилипают к глине и образуют глино-гумусовый комплекс. Также как и глина, гумус обладает большой поверхностью частиц и высокой катионообменной способностью. Эта способность особенно важна для почв с низким содержанием глины. Анионы в гумусе – это карбоксильные и фенольные группы. Благодаря своим химическим и физическим свойствам, гумус улучшает структуру почвы и ее аэрацию, а также повышает способность удерживать воду и питательные вещества.
Климатические факторы.
Свет
Свет необходим для жизни, так как это источник энергии для фотосинтеза, однако есть и другие аспекты его воздействия на живые организмы. Интенсивность света, его качество (длина волны, или цвет) и продолжительность освещения (фотопериод) могут оказывать различное влияние.
Необходимость света для растений существенно влияет на структуру сообществ. Распространение водных растений ограничено поверхностными слоями воды. В наземных экосистемах в процессе конкуренции за свет у растений выработались определенные стратегии, например быстрый рост в высоту, использование других растений в качестве опоры, увеличение поверхности листьев.
Температура
Главным источником тепла является солнечное излучение; им могут быть также геотермальные источники, но они играют важную роль только в немногих местообитаниях.
Температура, так же как интенсивность света, в большой мере зависит от географической широты, сезона, времени суток и экспозиции склона. Однако часто встречаются и узколокальные различия в температуре; это в особенности касается микроместообитаний, обладающих собственным микроклиматом. Растительность тоже оказывает некоторое влияние на температуру. Например, иная температура бывает под пологом леса или в меньшей степени внутри отдельных групп растений, а также под листьями отдельного растения.
Влажность и соленость.
Вода необходима для жизни и может быть важным лимитирующим фактором в наземных экосистемах. Вода поступает из атмосферы в виде осадков: дождя, снега, дождя со снегом, града или росы. В природе происходит непрерывный круговорот воды – гидрологический цикл, от которого зависит распределение ее на поверхности суши. Наземные растения поглощают воду главным образом из почвы. Быстрый дренаж, небольшое количество осадков из почвы, сильное испарение или сочетание всех этих факторов могут приводить к иссушению почв, а при изобилии, напротив, возможно их постоянное переувлажнение. Таким образом, количество воды в почве зависит от водоудерживающей способности самой почвы и от баланса между количеством выпадающих осадков и совместным результатом испарения и транспирации. Испарение происходит как с поверхности влажной растительности, так и с поверхности почвы.
Рациональное использование экосистем.
«Получение урожая» означает изъятие из экосистемы тех организмов или их частей, которые используются в пищу (или для иных целей). При этом желательно, чтобы экосистема производила пригодную для пищи продукцию наиболее эффективно. Это может быть достигнуто путем повышения урожайности культуры, уменьшения заболеваемости и помех со стороны других организмов или использования культуры, более приспособленной к условиям данной экосистемы.
Изучая продуктивность экосистем, мы имеем дело с потоком энергии, проходящих через ту или иную экосистему. Энергия поступает в биотический компонент экосистемы первичных продуцентов. Скорость накопления энергии первичными продуцентами в форме органического вещества, которое может быть использовано в пищу, называется первичной продукцией. Это важный параметр, так как им определяется общий поток энергии через биотический компонент экосистемы, а значит, и количество (биомасса) животных организмов, которые могут существовать в экосистеме.